Նորարարական նախագծերի տոնավաճառ Նորարարություններ մասնագիտական կրթության մեջ
Ցանկացած Փառատոնի ժողով Սպորտային միջոցառում Շնորհանդես Հանդիպում Կոնգրես Համերգ Իրադարձություն Մրցույթներ Ցուցահանդես-կոնֆերանս...
Քարտեզագրումը կոնտիգները կցելու ամենահեշտ ձևն է:
Գենոմի քարտեզագրումը կարող է լինել գենետիկ կամ ֆիզիկական:
Գենետիկական քարտեզագրում.
Պարունակում է գեն՝ ֆենոտիպային մարկեր;
Մոլեկուլային մարկերներ.
RFLP-ը (սահմանափակող հատվածի երկարության պոլիմորֆիզմ) գենոմային ԴՆԹ-ի ուսումնասիրության մեթոդ է՝ ԴՆԹ-ն կտրելով՝ օգտագործելով սահմանափակող էնդոնուկլեազներ (նուկլեինաթթուները մեջտեղում կտրել) և առաջացած բեկորները (սահմանափակումները) հետագա վերլուծելով գելային էլեկտրոֆորեզով։
SSLP (պարզ կրկնվող երկարության պոլիմորֆիզմ) - Կան 2 տեսակ՝ մինիարբանյակներ և միկրոարբանյակներ: Միկրոարբանյակներ– կրկնել 2,3,4 n.p. Սրանք ԴՆԹ պոլիմերազի աշխատանքի սխալներ են: Երբ կա մի տեղ, որտեղ օլիգոնուկլեոտիդը բազմիցս կրկնվում է, ԴՆԹ պոլիմերազի սայթաքման հավանականություն կա: Որոշակի հավանականության դեպքում այն կարող է անջատվել մատրիցից և միանալ հարևան կրկնությանը: Արդյունքում կրկնօրինակումը կամ կունենա մեկ ավել կամ մեկ պակաս կրկնություն.
SNP ( Մեկ նուկլեոտիդային պոլիմորֆիզմ) – 1 նուկլեոտիդի ԴՆԹ-ի հաջորդականության տարբերությունները նույն տեսակի ներկայացուցչի գենոմում.
Մոլեկուլային մարկերների հայտնաբերում.
1. ԴՆԹ հիբրիդացում:
1.1. Զոնդերը կզուգակցվեն միաշղթա ԴՆԹ-ի հետ՝ առաջացնելով ամբողջական կամ թերի լրացումներ:
1.2. Էլեկտրոֆորեզ PAA-ում. Տեղափոխում նիտրոցելյուլոզային թղթի միջոցով: Ռադիոպիտակավորված զոնդերը հիբրիդացվում են ԴՆԹ-ի հետ: Հայտնաբերվել է ավտոռադիոգրաֆիկ եղանակով:
Օգտագործվում է մոլեկուլային մարկերներ հայտնաբերելու համար.
1. Կրկնվող տեղորոշիչներ;

2. Սահմանափակող հատվածի պոլիմորֆիզմ (RFLP, RFLP) – սահմանափակող ֆերմենտի մշակում:



Կրկնվող տեսակները.
1. Կարճ (2-4 nt) պարզ հաջորդականության երկարության պոլիմորֆիզմ (SSLP);

2. Մինիարբանյակներ (մինչև 25 նտ.) VNTR;
3. Միկրարբանյակներ (դի- կամ տետրանուկլեոտիդներ) STR, SSR;
Ագարոզա գել մեկ նուկլեոտիդային պոլիմորֆիզմ (SNP);
(AGTCA Գ AAATC);
(AGTCA Գ AAATC);


ԴՆԹ-ի չիպերի օգտագործումը ֆոտոլիտոավտոգրաֆիայի միջոցով:
Ապակե սուբստրատը մշակվում է և դրա վրա կիրառվում է նուկլեոտիդ։ 5' ծայրը արգելափակված է, և լազերային լուսավորությունը հեռացնում է արգելափակող մասը: Այս կերպ կարելի է կիրառել հարյուր հազարավոր զոնդեր։

Ոչ հիբրիդացնող բեկորները լվանում են. որոշվում է բծերով (լյումինեսցենտով պիտակավորված ԴՆԹ-ից):

Թերություններ:
1. Սահմանափակ թույլատրելիություն (առավելագույնը 1 գեն 3,3 կբ-ին E. coli-ում և 1 գեն 10 կբ-ում S. cerevisiae-ում);
2. Սահմանափակ լուծում՝ անհավասար հատման պատճառով:
Ֆիզիկական քարտեզագրման հիմնական մեթոդները.
1. Սահմանափակման քարտեզագրում;
2. FiSH (լյումինեսցենտ in situ հիբրիդացում);
3. ՀՊԾ քարտեզագրում;
Սահմանափակման քարտեզագրումը գենոմային քարտեզի վրա հարաբերական վայրերի և սահմանափակման հատուկ վայրերի և դրանց միջև եղած հեռավորությունների որոշումն է:
Մեծ հողամասերի առանձնահատկությունները.
1. 7-8 նուկլեոտիդներ ճանաչող սահմանափակող ֆերմենտների օգտագործում;
2. սահմանափակող ֆերմենտների օգտագործումը, որոնց ճանաչման վայրը ներառում է նուկլեոտիդների հազվագյուտ համակցություններ.
3. Գելային էլեկտրոֆորեզի հատուկ տեխնիկայի կիրառում փոփոխական էլեկտրական դաշտով ԴՆԹ-ի մեծ բեկորները առանձնացնելու համար:
Դասական մոտեցումը էլեկտրոֆորեզն է՝ բեկորների չափը որոշելու համար: Անհրաժեշտ է առնվազն 2 սահմանափակող ֆերմենտ, որոնցից յուրաքանչյուրը մշակում է ուսումնասիրվող ԴՆԹ-ի հատվածը։
Առաջին բանը, որ դուք պետք է անեք, ընտրեք սահմանափակող ֆերմենտ, որն ավելի քիչ է կտրում, որպեսզի ստանաք ողջամիտ քանակությամբ բեկորներ: Ահա վիճակագրորեն հաշվարկված տեղանքի հաճախականությունը տարբեր էնդոնուկլեազների համար՝ տարբեր տեղամասերի չափսերով: Վերցվում են սահմանափակող ֆերմենտներ, որոնք ունեն ամենաերկար տեղամասը, գումարած սահմանափակող ֆերմենտները, որոնք ունեն նուկլեոտիդների հազվադեպ համակցություններ, հատուկ ընտրված են տեղամասերում: Այս կերպ մենք ստանում ենք սահմանափակման ռեակցիա խելամիտ թվով բեկորներով, բայց մենք դեռ պետք է որոշենք այդ բեկորների չափերը: Մեծ բեկորների չափը որոշելու համար օգտագործվում են հատուկ տեխնիկա՝ կա՛մ էլեկտրոֆորեզ փոփոխական դաշտում, կա՛մ վիզուալացման այլ մեթոդներ, որոնք համակցված են օպտիկական քարտեզագրման անվան տակ։ .
Նախ, այս կերպ դուք կարող եք գտնել ֆերմենտներ, որոնք նույնիսկ ավելի քիչ են կտրում: Մասնավորապես, օգտագործելով մարդու գենոմի օրինակը, այս կայքը ցույց է տալիս, թե ինչու մենք քիչ CG համակցություններ ունենք: Մարդու գենոմում CG տառերը, որոնք գտնվում են միմյանց կողքին, հանդիպում են միայն խթանող շրջաններում։ Համապատասխանաբար, դրանք չկան գենոմի մեծ մասում: Ինչու է դա տեղի ունենում: Քանի որ ցիտոսինը մեթիլացված է, այսինքն. ավելացվում է մեթիլ խումբ: Ցիտոզին է եղել, պարզվել է, որ 5-մեթիլցիտոզին է։ Ազոտային հիմքերի բաղադրության մեջ գտնվող ցանկացած ամինո խումբ ունակ է հիդրոլիզի բավականին մեծ հավանականությամբ։ Հիդրոլիզից ստացված մնացորդը ամինախմբի փոխարինումն է օքսո խմբով, իսկ եթե ցիտոսինը փոխարինվում է, ապա ստանում ենք ուրացիլ, որը հավասար հիմք է և, համապատասխանաբար, կտրվում է վերանորոգման համակարգով, բայց եթե այն 5- է. մեթիլցիտոզին, այնուհետև մենք ստանում ենք թիմին, և սա օրինական հիմք է ԴՆԹ-ում: Համապատասխանաբար, եթե C տառը կրիտիկական չէ գենոմային հաջորդականության համապատասխան տեղում, ապա էվոլյուցիոն ճնշումը բավականին ծանր է՝ փոխարինելով C տառը T տառով: Էվոլյուցիոն ճնշումը ավելի մեծ կլինի կոդավորման հաջորդականության մեջ և խթանող հաջորդականության մեջ: , քանի որ մեթիլացված CG հաջորդականությունները կարևոր կարգավորիչ ազդանշան են, որը վերահսկում է. Սրանք այսպես կոչված CPGS իրավունքներն են, որտեղ դրանք կան, գրեթե անկասկած կլինի ակտիվացված կարգավորող տարածք: Այսպիսով, եթե խնդիրը հազվագյուտ սահմանափակող ֆերմենտներ ընտրելն է, ապա անհրաժեշտ կլինի ընտրել սահմանափակող ֆերմենտներ, որոնց տեղանքը ներառում է նուկլեոտիդների այս համակցությունները։ Տեսնենք, թե ինչ կլինի .
Տարբերակներից մեկը էլեկտրոֆորեզն է պուլսացիոն դաշտում: Ստանդարտ էլեկտրոֆորեզը շատ վատ է հարմարեցված ավելի քան 50 հազար bp ունեցող բեկորների համար: Նրանք բոլորը մի փունջ հավաքվում են գելի վերին մասում, չեն առանձնանում, իսկ ոմանք նույնիսկ մնում են անցքի մեջ։ Նրանց առանձնացնելու համար փոխվում են էլեկտրոֆորեզի պայմանները՝ երկու էլեկտրոդները փոխվում են 4-ի և փոփոխական լարում է կիրառվում նախ մի էլեկտրոդի, ապա մյուսի վրա։ Երբ լարումը կիրառվում է առաջին էլեկտրոդի վրա, ԴՆԹ-ի բեկորը շարժվում է շատ կարճ հեռավորության վրա և խրվում ագարոզայի ծակոտիներում։ Լարումը փոխվում է, հոսանք է մատակարարվում մյուս էլեկտրոդներին, ԴՆԹ-ի բեկորը դուրս է քաշվում այն բջիջներից, որոնց մեջ այն խրված է և մի փոքր շարժվում է, հետո ուղղությունը նորից փոխվում է։ Շարժումը զիգզագ է ստացվում։ Բայց քայլերը շատ կարճ են և գումարվում են գծային շարժման: Իրականում ստացվում է սովորական էլեկտրոֆորեզ, տարբերությունն այն է, որ խոշոր բեկորները հստակ կառանձնանան, քանի որ շարժունակությունը բոլորովին այլ է. Ոչ այնքան կոշտ կապված բեկորների զանգվածի հետ: Էլեկտրոֆորեզը ունի մեկ մեծ թերություն՝ դա ԴՆԹ-ի չափի ուղղակի չափում չէ: Մոլեկուլային մարկերը կանցնի հետքերից մեկի երկայնքով և կհամեմատվի վազքի երկարությունը՝ անուղղակի ցուցիչ:

Այն, ինչ կոչվում է օպտիկական քարտեզագրում, փոխարինում է էլեկտրոֆորեզին, այստեղ բեկորների չափը ուղղակիորեն չափվում է։ Դրա էությունը կայանում է նրանում, որ ԴՆԹ-ն ուղղվում է մի քանի եղանակներից մեկով, այն դուրս է քաշվում ոլորված գնդակից: Ավելին, այն ձգվում է լարվածության տակ։ Ավելին, ԴՆԹ-ի մոլեկուլի լարվածությունը մշակվում է սահմանափակող ֆերմենտի միջոցով, կտրելուց հետո մոլեկուլի ծայրերը հեռանում են կտրվածքի տեղից, այս ամենը կարելի է տեսնել մանրադիտակի տակ։ Այնուհետև նման պատրաստուկը հետազոտվում է մանրադիտակի տակ, ԴՆԹ-ն ներկվում է ինտերկալատիվ լյումինեսցենտային ներկով և տեսանելի են կոտրման վայրերը։ Ուղղակի երկարության չափում մանրադիտակի տակ: Գծային լարված ԴՆԹ-ի ստացման երկու տարբերակ՝ երկու դեպքում էլ օգտագործվում է տարբեր փուլերի սահմանի հատկությունը։ Երբ ԴՆԹ-ն անցնում է փուլային միջերեսով, այն երկարանում է: Առաջին տեխնիկան օգտագործում է ԴՆԹ հալված ագարոզայի լուծույթում: ԴՆԹ-ն կիրառվում է ապակե սլայդի վրա, իսկ որոշ ժամանակ անց ագարոզը սկսում է կարծրանալ։ Կարծրացումը երբեք հավասարապես չի առաջանում, այն միշտ սկսվում է ինչ-որ տեղից: Ագարոզայի բյուրեղացման ճակատը շարժվում է սկզբնական կետից և տարածվում։ Երբ այն անցնում է ԴՆԹ-ի մոլեկուլով, իր հետ քաշում է այս մոլեկուլը և արդյունքում ստանում ենք լարված վիճակում գտնվող մոլեկուլ։ Այնուհետև այստեղ ավելացվում է ֆերմենտ, որոշ դեպքերում ֆերմենտը անմիջապես անշարժացվում է ապակու վրա, իսկ հետո համապատասխան բուֆերի հետ ավելացնում են մագնեզիումի իոններ, կամ գուցե հակառակը, հետո ֆերմենտը։ Բայց առաջին տարբերակն ավելի լավն է՝ ավելի լավ է ցրվում։ Մեկ այլ մեթոդ հիմնված է այն փաստի վրա, որ ծածկույթը շատ դանդաղ է հեռացվում ԴՆԹ լուծույթից։ Այն մշակվում է սիլանի լուծույթով, որը մեծացնում է ԴՆԹ-ի կլանումը, մոլեկուլների մի մասը ներծծվում է ապակու այս կտորի վրա և այն շատ դանդաղ, ժամում մի քանի միլիմետր, դուրս է քաշվում լուծույթից և երբ այն անցնում է Մակերեւութային լարվածության թաղանթ, ԴՆԹ-ի մոլեկուլները ցած են քաշվում այն վայրից, որտեղ ամրագրված են: Ուղղությունը հակառակ է ապակու շարժմանը: Մենք ստանում ենք լարված վիճակում գտնվող մոլեկուլ, ապա ապակին մշակվում է սահմանափակող ֆերմենտով:

Չզույգված վիճակում հիբրիդացում չկա: SNP-ների հայտնաբերումը լուծույթի հիբրիդացման միջոցով: Հիբրիդացման ժամանակ վարսահարդարիչը բացվում է։
Հաջորդ քարտեզագրման տեխնիկան լյումինեսցենտային հիբրիդացումն է պատրաստուկների վրա՝ FISH .
Այստեղ ցիտոլոգիական ներկով ներկված քրոմոսոմների վրա (ոլորված քրոմոսոմներ) ներկված լյուորեսցենտային զոնդի դիրքին համապատասխան տեղանքը ուղղակիորեն պատկերվում է: Դեղը կիրառվում է ապակու վրա, ամրացվում դրա վրա և ենթարկվում մասնակի դենատուրացիայի: Արդյունքում մենք ստանում ենք քրոմոսոմի երկարությամբ հատվածներ միաշղթա վիճակում։ Ավելացվում է նմուշ, որը հիբրիդացվում է այս տարածքների հետ, իսկ հետո նման սարքը ներկվում է համապատասխան ներկերով և մանրադիտակով: Մանրադիտակն իրականացվում է տեսանելի տիրույթում, լյումինեսցենտային մանրադիտակով: Նմուշը լազերով ճառագայթվում է որոշակի ալիքի երկարությամբ և նկատվում է ֆլյուորեսցենտ: Արդյունքում մենք ստանում ենք ֆլյուորեսցենտային օրինաչափություն, որը վերադրվում է տվյալ քրոմոսոմին բնորոշ ժապավենային օրինաչափությանը: Մենք հատուկ գենի համար զոնդ արեցինք և անմիջապես տեսանք, թե այս գենը որտեղ է գտնվում քրոմոսոմի վրա:
Թերությունները. բանաձեւը բավականին ցածր է, ոչ ավելի, քան 1 միլիոն ն: հարակից գոտիների միջև. Տեխնիկայի փոփոխություններն օգտագործվում են լուծումը բարձրացնելու համար:
Ամենապարզը իրականում ոչինչ չի փոխում՝ քրոմոսոմների մեխանիկական ձգումը: Քրոմոսոմային պատրաստուկը պտտվում է ցենտրիֆուգում ցածր արագությամբ: Քրոմոսոմները ձգվում են կենտրոնախույս ուժերի ազդեցությամբ, գլխավորը հեղափոխություններով չչափազանցելն է։ Սպիտակուցները չեն միանում և մնում են այն դիրքերում, որտեղ ամրացված են և կարող են գունավորվել։ Բանաձեւը մեծանում է 4-5-ով։ Մինչև 200 հազ. հարակից մարկերների միջև: Եթե ավելի բարձր լուծաչափ է անհրաժեշտ, ապա օգտագործվում են այլ փոփոխություններ՝ ոչ մետաֆազային քրոմոսոմներ (մինչև 20-25 հազար bp լուծաչափ) և մանրաթել-FISH, նույն քրոմոսոմները, որոնք միայն լրացուցիչ ձգվում են՝ ձգվում են գելում, մոլեկուլային «սանրում», բանաձեւը մինչեւ 10 հազ. n. Այս երկու մեթոդների թերությունն այն է, որ մենք չենք կարող կապել լյումինեսցենտային զոնդը բջջաբանական քարտեզի հետ: Մենք կարող ենք տեղադրել միայն 2 զոնդերը միմյանց նկատմամբ և որոշել զոնդերի միջև հեռավորությունը: Պետք է լինի մի հետաքննություն, որի դիրքորոշումը հայտնի է, և մենք պետք է միաժամանակ տեսնենք և՛ այս հետաքննությունը, և՛ մեզ հետաքրքրող հետաքննությունը։


Քարտեզագրման մեկ այլ մեթոդ է STS քարտեզագրումը: . Այն ինչ-որ չափով նման է գենետիկական քարտեզագրմանը, սակայն ռեկոմբինացիա օգտագործելու փոխարեն օգտագործում է հարաբերական հեռավորությունը որոշելու այլ եղանակ: Չափանիշն այստեղ ԴՆԹ-ի մասնատման հաճախականությունն է գենոմային գրադարանների և բեկորների կառուցման ժամանակ: Իրականում էությունը նույնն է՝ ԴՆԹ-ի մոլեկուլը կոտրվում է ցածր ուլտրաձայնի ազդեցությամբ կամ սահմանափակող ֆերմենտների ազդեցությամբ։ տարբերությունը նաև նրանում է, թե ինչ է ծառայում որպես մարկեր: STS մարկերները հայտնի ԴՆԹ-ի բեկորներ են, որոնք հայտնվում են մեկ անգամ գենոմում, և հաջորդականությունը պետք է հայտնի լինի:
Երբ հայտնի է նման հաջորդականությունը, այս 100-500 նտ-ի սահմաններում կարելի է ընտրել զույգ այբբենարան: և այլն և ստացեք PCR հատված, որով սովորաբար հայտնաբերվում է նման մարկեր: Այս մարկերները ստանալու համար հաջորդականություններ ընտրելու ամենապարզ ձևը EST հաջորդականությունների օգտագործումն է՝ cDNA-ի կարճ կտորներ, որոնք պատրաստված են սուրհանդակային ՌՆԹ-ից: Քանի որ դա mRNA-ի պատճենն է, երաշխավորված է, որ այն մեկ անգամ կհայտնվի գենոմում, քանի որ գեների մեծ մասը եզակի է: Ըստ այդմ, սա ի սկզբանե յուրահատուկ հաջորդականություն է։ Այլ մարկերներ, ավելի քիչ հարմար, մարկերներ են, որոնք պոլիմորֆ են պարզ հաջորդականությամբ՝ մինիարբանյակներ կամ միկրոարբանյակներ: Դուք կարող եք նաև օգտագործել պատահական գենոմային հաջորդականություններ, բայց դրանք ավելի դժվար են, քանի որ անհրաժեշտ է հստակ ապացուցել, որ դրանք եզակի գենոմային հաջորդականություններ են։ Նման մարկերների դիրքը որոշելու համար անհրաժեշտ է մեկ այլ բաղադրիչ՝ քարտեզագրման ռեագենտ։ Սա ընդամենը ԴՆԹ-ի բեկորների հավաքածու է որոշակի գրադարանից: Որքան ավելի մոտ լինեն մարկերները միմյանց իրական քրոմոսոմային քարտեզի վրա, այնքան մեծ է հավանականությունը, որ դրանք կհայտնվեն ԴՆԹ-ի նույն հատվածում:
Մարկերները սերտորեն կապված են և գտնվում են մոտակայքում, ուստի դրանք հայտնվում են մեծ քանակությամբ բեկորների մեջ: Ավելի հեռու գտնվող մարկերներն ամենից հաճախ ընկնում են ԴՆԹ-ի տարբեր բեկորների վրա: Եթե մարկերները նույնիսկ ավելի հեռու են, նրանց միջև հեռավորությունը կգերազանցի գրադարանի բեկորների չափը, և դրանք ընդհանրապես չեն հայտնաբերվի միասին: Այս ամենը վիճակագրորեն պոկված է. ելնելով գրադարանում այս մարկերների համատեղ առաջացման հաճախականությունից, հնարավոր է բավականին ճշգրիտ որոշել դրանց միջև հեռավորությունը իրական քրոմոսոմային քարտեզի վրա: Պայմանով, որ մասնատումը միայն պատահական է: Դրվագների աղբյուրները՝ ստանդարտ կլոնային գրադարան, ճառագայթային հիբրիդային բջիջների հավաքածու։



Ճառագայթային հիբրիդային բջիջները առավել հարմար են STS քարտեզագրման համար: Սա հին տեխնոլոգիա է, իրականում էուկարիոտական ԴՆԹ-ի in vivo կլոնավորման տարբերակ: Խոսքը ճառագայթված բջիջների հիբրիդացման մասին է չճառագայթված բջիջների հետ։ Ընդ որում, չճառագայթված բջիջները, որպես կանոն, չինական համստերի բջիջներ են, իսկ ճառագայթված բջիջները ցանկացած կաթնասունի բջիջներ են, այդ թվում. մարդ. Եթե ենթադրենք, որ դրանք մարդկային բջիջներ են, դրանք ենթարկվում են γ-ճառագայթման շատ մեծ չափաբաժինների, և այդպիսի բջիջը դառնում է ոչ կենսունակ, մենք ստանում ենք լուրջ մասնատված ԴՆԹ-ով բջիջ, և անհնար է վերականգնել նման քրոմոսոմները: Եթե նման բջիջը միաձուլվի մեկ այլ կաթնասունի բջիջի հետ, ապա անձեռնմխելի բջիջի վերականգնողական համակարգը կփորձի վերականգնել այս երկշղթայի ճեղքերը, և արդյունքը կլինի այս ԴՆԹ-ի բեկորների պատահական ընդգրկումները նրա անձեռնմխելի քրոմոսոմների տարբեր մասերում: Այն, ինչ հնարավոր չէր միացնել բջիջների բաժանման ժամանակ, կվերացվի, և այն, ինչ միացված է, որպես ստանդարտ կժառանգվի այս բջջային գծում: Քանի որ յուրաքանչյուր քրոմոսոմում ներառված են օտար ԴՆԹ-ի մի քանի բեկորներ, ճառագայթման ճիշտ չափաբաժնով, այդպիսի հիբրիդոմների հարյուր տողից մոտ 100 բջիջները բավարար են էուկարիոտիկ գենոմի մանրամասն STS քարտեզ կառուցելու համար:
PCR-ի միջոցով գրանցվում է ԴՆԹ-ի մեկ նմուշում մարկերների առկայությունը և հաշվարկվում է այդ մարկերների հաճախականությունը բոլոր գծային տարբերակներում: Կան տարբեր ավտոմատացման մեխանիզմներ.
Դրա էությունը հանգում է նրան, որ հայտնաբերման համար օգտագործվում է ոչ թե էլեկտրոֆորեզ, այլ լյումինեսցենտային պիտակ, և այս պիտակը այստեղ ներառված է բավականին խելացի ձևով։ Ռեակցիային ավելացվում են լյումինեսցենտային նուկլեոտիդներ, սակայն հայտնաբերման համակարգը ի վիճակի է հայտնաբերել միայն բևեռացված ազդանշան, սկզբում ազդանշանը բևեռացված չէ. հայտնաբերում չի լինում: Իրավիճակը սկսվում է, երբ նման նուկլեոտիդն ընդգրկվում է ԴՆԹ-ում։ Այս նուկլեոտիդները տերմինատոր են, ուստի դրանք միացված են միայն մեկ անգամ և մի փոքր այլ կերպ ֆլյուորեսվում են միացնելուց հետո: Կատարվում է PCR, եթե արտադրանքը առկա է, ապա ավելացվում է մեկ այլ ներքին այբբենարան; այն հիբրիդացվում է ապրանքային շղթաներից մեկի հետ և դրան ավելացվում է այս նուկլեոտիդը: Դրանից հետո ազդանշանը դառնում է բևեռացված և հնարավոր է հայտնաբերել: Եթե ապրանք չկա, ազդանշան չի լինի։ PCR հայտնաբերման այս փոփոխությամբ հնարավոր է արագ կառուցել գենոմի ֆիզիկական քարտեզը էուկարիոտիկ գենոմի մասշտաբով:
Երբ մենք սկսեցինք խոսել sts քարտեզագրման մասին, ես ասացի, որ քարտեզագրման համար բեկորների աղբյուրի երկրորդ տարբերակը միայն ստանդարտ կլոնային գրադարանն է։ Դա մի փոքր ավելի քիչ հարմար է այն առումով, որ շատ ավելի շատ կլոններ պետք է օգտագործվեն, մեկ հատվածի վրա համակցված դեպքերի հաճախականությունը ավելի ցածր է, բայց զննումն ավելի ընդարձակ է: Բայց կան առավելություններ, որոնք գոնե մասամբ փոխհատուցում են այս թերությունը, դրանք այն են, որ նույն կլոնները կարող են օգտագործվել քարտեզը կառուցելու համար, և նույն կլոնները կարող են այնուհետև վերցնել հաջորդականության համար: Նույն կլոնային գրադարանը կարող է օգտագործվել քարտեզագրման, այնուհետև նուկլեոտիդների հաջորդականությունը որոշելու համար, ուստի հատուկ որևէ բան նախագծելու կարիք չկա, և արդեն գոյություն ունեցող կլոնային գրադարանը կարող է օգտագործվել նման քարտեզագրման համար։




Դե, երբ կար բջջաբանություն, ձեզ երևի ասվել է ֆլյուորեսցենտային օժանդակությամբ բջիջների տեսակավորման սկզբունքի մասին, այսինքն՝ այս բջիջները ներկված են լյումինեսցենտային ներկով, այսինքն՝ այս տեխնիկայի շատ աննշան փոփոխությունը թույլ է տալիս բացել ոչ բջիջները, բայց քրոմոսոմներ. Եվ այս սկզբունքը, համապատասխանաբար, հնարավորություն է տալիս ամբողջ քրոմոսոմային հավաքածուն բաժանել առանձին քրոմոսոմների, այնուհետև մեկ քրոմոսոմի ԴՆԹ-ից գրադարաններ են պատրաստում։ Սա հսկայական օգնություն է, քանի որ մի բան է աշխատել 3 միլիարդ նուկլեոտիդային զույգ գենոմի մասշտաբով, մեկ այլ բան՝ մեկ քրոմոսոմի մասշտաբով, այնուհետև մենք ստանում ենք միջինը 150 միլիոն: Դուք զգում եք, թե ինչպես է սանդղակը նվազում և բնականաբար սա մեթոդը օգտագործվում է. Հուսով եմ, որ դուք լավ հիշում եք սա, այստեղ ամեն ինչ չափազանց պարզ է: Այն, ինչ պետք է տեսակավորվի, ներկված է լյումինեսցենտային ներկով, այս դեպքում՝ քրոմոսոմներով։ Ներկանյութի քանակը, որը կապվում է քրոմոսոմին, կախված է մի քանի գործոններից, առաջին հերթին, քրոմոսոմի չափից, սա հիմնական գործոնն է, և երկրորդը, սպիտակուցների կառուցվածքը, որոնք կապված են այս քրոմոսոմի հետ: Սպիտակուցները մի փոքր տարբերվում են, հիմնականում տարբերվում են հետերոքրոմատինի քանակով, լավ, սա որոշում է, որ քրոմոսոմի չափի և ներկման ինտենսիվության միջև ուղղակի կապ չկա: Դե ինչ է լինում հետո, հետո լուծույթը կաթում է փորձանոթից փոքր կաթիլներով, յուրաքանչյուր կոնցենտրացիան ընտրվում է այնպես, որ կաթիլը կա՛մ դատարկ է, կա՛մ պարունակում է մեկ քրոմոսոմ, ոչ մի դեպքում երկու։ Այնուհետև կաթիլը կողքից լուսավորվում է լազերային ճառագայթով, և եթե կաթիլում առկա է քրոմոսոմ, ֆլյուորեսցենցիան գրգռվում է, և ֆլյուորեսցենտը հայտնաբերվում է դետեկտորի միջոցով: Լյումինեսցենցիայի ինտենսիվությունը համաչափ է կապած ներկի քանակին: Այս քանակությունը յուրովի է բնութագրում յուրաքանչյուր քրոմոսոմին, այս փուլում կարելի է ասել, թե որ քրոմոսոմն է գտնվում այս կաթիլում։ Այնուհետև այս զույգ էլեկտրոդները, ֆլյուորեսցենտային ինտենսիվությանը համապատասխան, լիցքավորում են կաթիլը, որպեսզի ստացվի բավական հզոր էլեկտրական դաշտ։ Որքան մեծ է ֆլյուորեսցենտը, այնքան ավելի շատ լիցք է փոխանցվում կաթիլին: Կաթիլն ավելի է թռչում, անցնելով հաջորդ զույգ էլեկտրոդների միջով: Այս էլեկտրոդները կաթիլը շեղում են աջ կամ ձախ՝ ըստ լիցքի։ Որքան մեծ է լիցքը, այնքան մեծ է շեղումը մի ուղղությամբ, ընդհանուր առմամբ այն կշեղվի մեկ ուղղությամբ և, համապատասխանաբար, յուրաքանչյուր քրոմոսոմ կաթելու է իր փորձանոթի մեջ։ Մարդկային քրոմոսոմների դեպքում ներկանյութի ճիշտ ընտրությունը հնարավորություն է տալիս բոլոր քրոմոսոմները, բացառությամբ մեկ զույգի, առանձնացնել առանձին փորձանոթների մեջ։ Մնացած զույգ քրոմոսոմների համար վերցվում է մեկ այլ ներկ, որը մի փոքր այլ կերպ է կապվում, և այս զույգը նույնպես կարելի է առանձնացնել։ Այսպիսով, մենք ստանում ենք առանձին քրոմոսոմների պատրաստուկներ ընդհանուր ԴՆԹ պատրաստումից, ինչը զգալիորեն հեշտացնում է հետագա աշխատանքը։ Այստեղ մենք ավարտում ենք քարտեզագրումը և գրադարանները:
Գենետիկ քարտեզագրումը դեռ օգտագործվում է և կշարունակվի կիրառվել, քանի որ... Բացի գենոմի քարտեզները կազմելուց, այն ունի կարևոր կիրառություն՝ կա մարկերի օգնությամբ ընտրության տեխնիկա, որը թույլ է տալիս շատ արագ ներկայացնել ցանկալի գենը:
Վայրի բնության նման նոր սորտի գենոտիպը հստակորեն վերահսկում է այս գենի առկայությունը մոլեկուլային մարկերով: Թերությունը սահմանափակ ճշգրտությունն է: E. coli-ի և խմորիչի համար գենետիկական քարտեզագրումը շարունակվում է 50-ականներից և քարտեզագրվել է հազար գեն: E.coli-ի և խմորիչի դեպքում նման քարտեզների առավելագույն խտությունը մեկ գեն է 3 հազար bp-ում։ E. coli-ում և մեկ գեն 10 հազար bp-ում: խմորիչի մեջ։ Երկու դեպքում էլ ստացվում է, որ հնարավոր է քարտեզագրել միայն յուրաքանչյուր երրորդ, ամեն չորրորդ գենը. Ավելի մեծ ճշգրտություն հնարավոր չէ հասնել: Երկրորդ խնդիրը կապված է այն փաստի հետ, որ հատումը տեղի է ունենում անհավասարաչափ քրոմոսոմի երկարությամբ, կան հատման թեժ կետեր, կան քրոմոսոմի հատվածներ, որոնք արգելափակում են հատումը. սա լրացուցիչ սխալ է մտցնում և, բացի այդ, կարող է փոխել որոշ տեղանքների դիրքերը գենետիկ քարտեզի վրա: Նաեւ անզգուշության պատճառով ավելանում է փորձարարական սխալների թիվը։
Քարտեզագրումն ունի 2 առաջադրանք. որոշել քրոմոսոմի վրա որոշակի շրջանների հարաբերական դիրքը, իսկ երկրորդ խնդիրն է որոշել նրանց միջև եղած հեռավորությունը. Նրանք. Ինչ վերաբերում է հարաբերական դիրքին, ապա գեների քարտեզագրումը քիչ թե շատ դրանով է հաղթահարում, եթե նայեք սլայդին, ապա այս մարկերների դիրքերը համընկնում են, բացառությամբ երկուսի, դրանք փոխված են, ակնհայտորեն այստեղ ինչ-որ անոմալիա կա: Ինչ վերաբերում է մարկերների միջև եղած հեռավորությանը, ապա այստեղ ամեն ինչ մի փոքր ավելի վատ է. տեսնում եք, որ որոշ հեռավորություններ պարզվում են մի փոքր ավելի մեծ, ոմանք ավելի փոքր, քան իրական հեռավորությունները: Համապատասխանաբար, այժմ ավելի ու ավելի կարևոր են դառնում ֆիզիկական քարտեզագրման տեխնիկան, որոնք այժմ գենոմային նախագծի անբաժանելի մասն են կազմում ինչպես դասական, այնպես էլ որսորդական հրացանի մոտեցման մեջ, քանի որ շատ դեպքերում ավարտական փուլում բացերը վերացնելու համար դուք դեռ չի կարող անել առանց քարտեզի, հատկապես էուկարիոտների գենոմների համար:
ԱԼ-ՖԱՐԱԲԻԻ ԱՆՎԱՆ ՂԱԶԱԽԻ ԱԶԳԱՅԻՆ ՀԱՄԱԼՍԱՐԱՆ
Ֆակուլտետը: կենսաբանություն և կենսատեխնոլոգիա
բաժին: կենսատեխնոլոգիա
«ՎԵՐացական»
Թեմայի շուրջ. ԳԵՆԵՏԻԿ ԿԱՊԸ ԵՎ ՄԱՐԴԿԱՅԻՆ ԳԵՆԵՐԻ ՔԱՐՏԵԶԱԳՐՈՒՄԸ.
Ավարտված : 3-րդ կուրսի ուսանողներ (med.bt.)
Նուրալիբեկով Ս.Շ.
Դավրոնովա Մ.Ա.
Ես ստուգել եմ : բ.գ.թ. , ամբիոնի դոցենտմոլեկուլային
կենսաբանություն և գենետիկաՕմիրբեկովա Ն.Ժ.
ԱԼՄԱԹԻ 2018
Գենետիկական կապի քարտեզներ……………………………………………………………………..3
Գենետիկական կապի քարտեզների կառուցման ժամանակակից մեթոդներ……………………..5
PCR-ն մարդու գենոմի հետազոտության մեջ……………………………………………………8
Ցածր լուծաչափով ֆիզիկական քարտեզներ…………………………………………………..9
Բարձր լուծաչափով ֆիզիկական քարտեզներ………………………………………………………………11
Օգտագործված աղբյուրների ցանկ……………………………………………………………….13
Մարդու գենոմի առաջնային կառուցվածքի քարտեզագրում և որոշում
Մոլեկուլային գենետիկայի մեջ առավել հաճախ օգտագործվող գեների կառուցվածքն ու մեխանիզմները ուսումնասիրելու հիմնական մեթոդների համառոտ վերանայումից հետո տեղին է թվում, օգտագործելով մարդու գենոմի օրինակը, ավելի մոտիկից նայել այդ մեթոդների և դրանց գործնական կիրառմանը: փոփոխություններ մեծ գենոմների ուսումնասիրության համար: Մարդու գենոմը, գենետիկական տեղեկատվության այս հսկայական շտեմարանը համակողմանի ուսումնասիրելու նպատակով վերջերս մշակվել և իրականացվում է «Մարդու գենոմի նախագիծ» միջազգային հատուկ ծրագիրը։ Ծրագրի հիմնական նպատակն է կառուցել մարդկային 24 քրոմոսոմներից յուրաքանչյուրի համապարփակ, բարձր լուծաչափով գենետիկական քարտեզներ, որոնք, ի վերջո, պետք է ավարտվեն այս քրոմոսոմների ԴՆԹ-ի ամբողջական առաջնային կառուցվածքի որոշմամբ: Ներկայումս նախագծի վրա աշխատանքները բուռն ընթացքի մեջ են։ Եթե այն հաջողությամբ ավարտվի (և նախատեսվում է դա տեղի ունենալ 2003 թվականին), մարդկությունը հեռանկարներ կունենա իր յուրաքանչյուր գենի ֆունկցիոնալ նշանակության և գործելու մեխանիզմների, ինչպես նաև մարդու կենսաբանությունը կառավարող գենետիկական մեխանիզմների մանրակրկիտ ուսումնասիրության համար, և պարզել իր մարմնի պաթոլոգիական վիճակների մեծ մասի պատճառները:
Մարդու գենոմի քարտեզագրման հիմնական մոտեցումները
Մարդու գենոմի ծրագրի հիմնական առաջադրանքի լուծումը ներառում է երեք հիմնական փուլ. Առաջին փուլում անհրաժեշտ է հատուկ բաժանել յուրաքանչյուր առանձին քրոմոսոմ ավելի փոքր մասերի` թույլ տալով դրանց հետագա վերլուծությունը՝ օգտագործելով հայտնի մեթոդները։ Հետազոտության երկրորդ փուլը ներառում է ԴՆԹ-ի այս առանձին բեկորների հարաբերական դիրքի որոշումը միմյանց նկատմամբ և դրանց տեղայնացումը հենց քրոմոսոմներում: Վերջնական փուլում անհրաժեշտ է իրականում որոշել քրոմոսոմի յուրաքանչյուր բնորոշ բեկորների առաջնային ԴՆԹ կառուցվածքը և կազմել դրանց նուկլեոտիդների ամբողջական շարունակական հաջորդականությունը: Խնդրի լուծումն ամբողջական չի լինի, եթե հնարավոր չլինի տեղայնացնել օրգանիզմի բոլոր գեները հայտնաբերված նուկլեոտիդային հաջորդականություններում եւ որոշել դրանց ֆունկցիոնալ նշանակությունը։ Վերոհիշյալ երեք փուլերի անցումը պահանջվում է ոչ միայն մարդու գենոմի համապարփակ բնութագրերը ստանալու համար, այլև ցանկացած այլ մեծ գենոմի:
Գենետիկական կապի քարտեզներ
Գենետիկական կապի քարտեզները առանձին քրոմոսոմների վրա գենետիկական մարկերների հարաբերական դիրքերի միաչափ դիագրամներ են: Գենետիկ մարկերները հասկացվում են որպես ցանկացած ժառանգական ֆենոտիպային հատկանիշ, որը տարբերվում է առանձին անհատների միջև: Գենետիկական մարկերների պահանջներին համապատասխանող ֆենոտիպային գծերը շատ բազմազան են։ Դրանք ներառում են ինչպես վարքագծային բնութագրեր կամ որոշակի հիվանդությունների հակվածություն, այնպես էլ ամբողջ օրգանիզմների կամ դրանց կառուցվածքով տարբերվող մակրոմոլեկուլների մորֆոլոգիական բնութագրերը: Կենսաբանական մակրոմոլեկուլների ուսումնասիրման պարզ և արդյունավետ մեթոդների մշակման հետ մեկտեղ նման հատկանիշները, որոնք հայտնի են որպես մոլեկուլային մարկերներ, առավել հաճախ օգտագործվում են գենետիկ կապի քարտեզների կառուցման մեջ: Նախքան նման քարտեզների կառուցման մեթոդները և գենոմի հետազոտության համար դրանց նշանակությունը դիտարկելը, անհրաժեշտ է հիշել, որ «կապ» տերմինը օգտագործվում է գենետիկայի մեջ՝ նշելու ծնողներից մեկից երկու հատկանիշների համատեղ փոխանցման հավանականությունը։ սերունդ.
Երբ մեյոզի փուլում կենդանիների և բույսերի մոտ ձևավորվում են սեռական բջիջներ (գամետներ), սովորաբար տեղի է ունենում հոմոլոգ քրոմոսոմների սինապսիս (կոնյուգացիա): Հոմոլոգ քրոմոսոմների քույր քրոմատիդներն իրենց ողջ երկարությամբ միացված են միմյանց, և խաչասերման արդյունքում (քրոմատիդների միջև գենետիկական վերահամակցում) դրանց մասերը փոխանակվում են։ Որքան հեռու են երկու գենետիկական մարկերները միմյանցից քրոմատիդում, այնքան մեծ է հավանականությունը, որ խաչմերուկի համար անհրաժեշտ քրոմատիդային ընդմիջումը տեղի կունենա նրանց միջև, և նոր գամետին պատկանող նոր քրոմոսոմի երկու մարկերները կառանձնանան յուրաքանչյուրից: այլ, այսինքն. դրանց կպչունությունը կխախտվի: Գենետիկ մարկերների կապող միավորը մորգանիդն է (Morgan unit, M), որը պարունակում է 100 ցենտիմորգանիդ (սմ): 1 սմ-ը համապատասխանում է գենետիկական քարտեզի վրա երկու մարկերների միջև եղած ֆիզիկական հեռավորությանը, որոնց միջև վերամիավորումը տեղի է ունենում 1% հաճախականությամբ: Հիմքերի զույգերով արտահայտված 1 սմ-ին համապատասխանում է 1 մլն bp։ (m.b.o.) ԴՆԹ.
Գենետիկական կապի քարտեզները ճիշտ արտացոլում են քրոմոսոմների վրա գենետիկական մարկերների հերթականությունը, սակայն դրանց միջև առաջացած հեռավորությունները չեն համապատասխանում իրական ֆիզիկական հեռավորություններին: Այս փաստը սովորաբար կապված է այն փաստի հետ, որ առանձին քրոմոսոմային հատվածներում քրոմատիդների միջև ռեկոմբինացիայի արդյունավետությունը կարող է շատ տարբեր լինել: Մասնավորապես, այն ճնշված է քրոմոսոմների հետերոխրոմատիկ շրջաններում։ Մյուս կողմից, քրոմոսոմներում հաճախ առաջանում են ռեկոմբինացիոն թեժ կետեր։ Ֆիզիկական գենետիկական քարտեզներ կառուցելու համար ռեկոմբինացիոն հաճախականությունների օգտագործումը՝ առանց այդ գործոնները հաշվի առնելու, կհանգեցնի գենետիկական մարկերների միջև իրական հեռավորությունների աղավաղումների (համապատասխանաբար թերագնահատում կամ գերագնահատում): Այսպիսով, գենետիկական կապի քարտեզները ամենաքիչ ճշգրիտն են առկա գենետիկական քարտեզների բոլոր տեսակներից և կարող են համարվել միայն որպես իրական ֆիզիկական քարտեզների առաջին մոտարկում: Այնուամենայնիվ, գործնականում հենց նրանք են և միայն նրանք, որոնք հնարավորություն են տալիս ուսումնասիրության առաջին փուլերում տեղայնացնել բարդ գենետիկական մարկերները (օրինակ, հիվանդության ախտանիշների հետ կապված) և հնարավոր դարձնել դրանց հետագա ուսումնասիրությունը: Պետք է հիշել, որ հատման բացակայության դեպքում առանձին քրոմոսոմի վրա տեղակայված բոլոր գեները միասին կփոխանցվեն ծնողներից սերունդներին, քանի որ դրանք ֆիզիկապես կապված են միմյանց հետ: Հետևաբար, առանձին քրոմոսոմները կազմում են գենային կապի խմբեր, և գենետիկ կապի քարտեզների կառուցման առաջին խնդիրներից մեկը հետազոտվող գենը կամ նուկլեոտիդային հաջորդականությունը որոշակի կապող խմբին վերագրելն է: Հաջորդը Աղյուսակում թվարկված են ժամանակակից մեթոդներ, որոնք, ըստ Վ.Ա. McKusick-ը ամենից հաճախ օգտագործվում էր գենետիկ կապի քարտեզներ կառուցելու համար մինչև 1990 թվականի վերջը:
Գենետիկ կապի քարտեզների կառուցման ժամանակակից մեթոդներ
| Մեթոդ | Քարտեզագրված տեղանքների քանակը |
| Սոմատիկ բջիջների հիբրիդացում | 1148 |
| In situ հիբրիդացում | 687 |
| Ընտանիք | 466 |
| Դոզայի ազդեցության որոշում | 159 |
| Սահմանափակման քարտեզագրում | 176 |
| Քրոմոսոմային շեղումների օգտագործումը | 123 |
| Սինթենիայի օգտագործումը | 110 |
| Ճառագայթման հետեւանքով առաջացած գեների տարանջատում | 18 |
| Այլ մեթոդներ | 143 |
| Ընդամենը | 3030 |
In situ հիբրիդացում. In situ հիբրիդացման մեթոդը լայնորեն կիրառվում է նաև քրոմոսոմների վրա նուկլեոտիդային հաջորդականությունների քարտեզագրման համար։ Այդ նպատակով ֆիքսված քրոմոսոմների պատրաստուկները հիբրիդացվում են (ինկուբացվում են բարձր ջերմաստիճանում, որին հաջորդում է սառեցումը) ուսումնասիրվող նուկլեոտիդային հաջորդականությունների հետ՝ պիտակավորված ռադիոակտիվ, լյումինեսցենտային կամ այլ պիտակով: Չկապված պիտակը լվանալուց հետո մնացած պիտակավորված նուկլեինաթթվի մոլեկուլները կապված են քրոմոսոմային շրջանների հետ, որոնք պարունակում են ուսումնասիրվող պիտակավորված նուկլեոտիդային հաջորդականություններին լրացնող հաջորդականություններ: Ստացված հիբրիդները վերլուծվում են մանրադիտակի միջոցով կամ ուղղակիորեն կամ ավտոռադիոգրաֆիայից հետո: Մեթոդների այս խումբը բնութագրվում է ավելի բարձր լուծաչափով, քան սոմատիկ բջիջների հիբրիդացումը, քանի որ դրանք թույլ են տալիս տեղայնացնել ուսումնասիրված նուկլեոտիդային հաջորդականությունները քրոմոսոմների վրա: Մարդկային գենոմի ծրագրի առաջընթացի հետ ավելի ու ավելի շատ մեկուսացված նուկլեոտիդային հաջորդականություններ հասանելի են դառնում հետազոտողների համար, որոնք կարող են օգտագործվել որպես in situ հիբրիդացման հետազոտողներ: Այս առումով վերջին ժամանակներս այս մեթոդները կիրառման հաճախականությամբ առաջին տեղն են զբաղեցրել։ Մեթոդների ամենատարածված խումբը կոչվում է fluorescence in situ հիբրիդացում (FISH), որն օգտագործում է լյումինեսցենտային պիտակ պարունակող պոլինուկլեոտիդային զոնդեր: Մասնավորապես, 1996 թվականին տպագրվել է ավելի քան 600 հոդված, որտեղ նկարագրված է այս մեթոդի կիրառումը:
Ընտանեկան գենետիկ կապի վերլուծություն. Մեթոդների այս խումբը հաճախ օգտագործվում է բժշկական գենետիկայի մեջ՝ բացահայտելու կապը (կապը) անհայտ գենի մուտացիայի հետևանքով առաջացած հիվանդության ախտանիշների և այլ գենետիկական մարկերների միջև: Այս դեպքում հիվանդության ախտանիշներն իրենք են հանդես գալիս որպես գենետիկական մարկերներից մեկը։ Մարդու գենոմում հայտնաբերվել են մեծ թվով պոլիմորֆիզմներ, ներառյալ RFLP-ները: RFLP-ները քիչ թե շատ հավասարաչափ բաշխված են մարդու գենոմում՝ միմյանցից 5–10 սմ հեռավորության վրա: Որքան ավելի մոտ են առանձին պոլիմորֆ լոկուսները տեղակայված հիվանդության համար պատասխանատու գենին, այնքան քիչ հավանական է, որ դրանք առանձնանան մեյոզի վերակոմբինացիայի ժամանակ, և ավելի հաճախ դրանք միասին կհայտնվեն հիվանդ անհատի մոտ և միասին կփոխանցվեն ծնողներից սերունդներին: Կլոնավորելով գենոմի ընդլայնված հատվածը, որը ներառում է համապատասխան պոլիմորֆ մարկեր (դրա ընտրությունը գենոմային ԴՆԹ գրադարանից կատարվում է զոնդի միջոցով), հնարավոր է դրա հետ միաժամանակ մեկուսացնել ժառանգական հիվանդություն առաջացնող գենը: Նման մոտեցումները, մասնավորապես, հաջողությամբ կիրառվել են Դյուշենի մկանային դիստրոֆիայի, երիկամների կիստիկական ֆիբրոզի և միոտոնիկ դիստրոֆիայի դեպքում ընտանեկան վերլուծության և համապատասխան գեների մեկուսացման համար: Մարդու գենոմի առանձին RFLP-ների տեղեկատվական բովանդակությունը կախված է հետազոտվող պոպուլյացիայի մեջ դրանց հետերոզիգոտության մակարդակից: RFLP-ի՝ որպես գենետիկական մարկերի ինֆորմատիվության չափանիշ, ըստ Դ. Բոտշտայնի և այլոց (1980) առաջարկի, համարվում է պոլիմորֆիզմի տեղեկատվական բովանդակության արժեքը՝ PIC (պոլիմորֆիզմի տեղեկատվական բովանդակություն), որը թվի հարաբերակցությունն է։ խաչեր, որոնցում ծնողներից առնվազն մեկի մոտ հետազոտվող պոլիմորֆ մարկերն ունի հետերոզիգոտ վիճակում, բոլոր խաչերի նկատմամբ:
Գենի չափաբաժնի ազդեցության որոշում և քրոմոսոմային շեղումների օգտագործում . Այս մեթոդները հայտնաբերում են փոխկապակցվածություն ուսումնասիրվող գենի արտահայտման մակարդակի և անուպլոիդ բջջային գծերում հատուկ քրոմոսոմների քանակի կամ քրոմոսոմների կառուցվածքային վերադասավորումների միջև (քրոմոսոմային մուտացիաներ - շեղումներ): Անեուպլոիդիան բջիջի, հյուսվածքի կամ ամբողջ օրգանիզմի առկայությունն է մի շարք քրոմոսոմներով, որոնք հավասար չեն տվյալ կենսաբանական տեսակի համար բնորոշին: Քրոմոսոմային շեղումները քրոմոսոմային շրջանների տեղափոխման տեսքով նույն կամ այլ քրոմոսոմների հետերոքրոմատիկ շրջաններ հաճախ ուղեկցվում են տեղափոխված շրջաններում կամ ընդունող քրոմոսոմում տեղակայված գեների տրանսկրիպցիայի ճնշմամբ (մոզաիկ դիրքի էֆեկտ):
Սինթենիայի օգտագործումը.Սինտենիան տարբեր կենսաբանական տեսակների օրգանիզմների գենային կապի խմբերի կառուցվածքային նմանությունն է: Մասնավորապես, մարդու և մկան գենոմում հայտնի են գեների մի քանի տասնյակ սինթենիկ խմբեր։ Սինթենիայի ֆենոմենի առկայությունը հնարավորություն է տալիս նեղացնել քրոմոսոմների վրա ուսումնասիրվող գենի տեղակայման որոնումը` սահմանափակելով այն կոնկրետ սինթենիկ խմբին պատկանող հայտնի գեների տարածաշրջանով:
Իոնացնող ճառագայթման արդյունքում առաջացած գեների տարանջատում: Օգտագործելով այս մեթոդը, ուսումնասիրվող գեների միջև հեռավորությունը որոշվում է իոնացնող ճառագայթման որոշակի ստանդարտ չափաբաժինով բջիջների ճառագայթումից հետո դրանց տարանջատման (տարանջատման) հավանականության գնահատմամբ: Ճառագայթված բջիջները փրկվում են մահից կրծողների սոմատիկ բջիջների հետ հիբրիդացման միջոցով, և որոշվում է ճառագայթահարված բջիջների ուսումնասիրված մարկերների առկայությունը սոմատիկ հիբրիդներում մշակույթի մեջ: Արդյունքում հնարավոր է եզրակացություն անել այս գեների միջև կապի (ֆիզիկական հեռավորության) առկայության կամ բացակայության մասին։
Ի թիվս այլ մեթոդներՊետք է նշել մեթոդները, որոնք հիմնված են գեների քարտեզագրման համար ԴՆԹ-ի մեծ բեկորների օգտագործման վրա, որոնք ձևավորվել են մեծ կտրվածքով սահմանափակող ֆերմենտների ազդեցության տակ: Գենոմային ԴՆԹ-ի տրոհումից հետո ստացված բեկորները էլեկտրոֆորեզի միջոցով բաժանվում են իմպուլսային էլեկտրական դաշտում, այնուհետև դրանք հարավային հիբրիդացվում են քարտեզագրված գեներին համապատասխան զոնդերով: Եթե հիբրիդացումից հետո երկու զոնդերի ազդանշանները տեղայնացված են ԴՆԹ-ի միևնույն մեծ հատվածի վրա, դա վկայում է նման գեների սերտ կապի մասին:
PCR մարդու գենոմի հետազոտության մեջ
Պոլիմերազային շղթայական ռեակցիան կենտրոնական տեղ է զբաղեցնում Մարդու գենոմի ծրագրի գործնական իրականացման մոտեցումների մշակման մեջ։ Ինչպես նշվեց վերևում, PCR-ը կարող է արագ և արդյունավետ կերպով ուժեղացնել մարդու գենոմի գրեթե ցանկացած կարճ հատված, և արդյունքում ստացված PCR արտադրանքը կարող է օգտագործվել որպես զոնդ՝ հարավային կամ in situ հիբրիդացման միջոցով քրոմոսոմների համապատասխան շրջանները քարտեզագրելու համար:
STS հայեցակարգ.Քննարկվող մարդկային գեների քարտեզագրման ծրագրի հիմքում ընկած հիմնական հասկացություններից մեկը հաջորդականությամբ պիտակավորված կայքերի (STS) հասկացությունն է: Համաձայն այս հայեցակարգի, ԴՆԹ-ի բոլոր բեկորները, որոնք օգտագործվում են գենետիկ կամ ֆիզիկական քարտեզներ կառուցելու համար, կարող են եզակիորեն նույնականացվել 200–500 bp նուկլեոտիդային հաջորդականությամբ, որը եզակի կլինի այդ հատվածի համար: Այս տեղամասերից յուրաքանչյուրը պետք է հաջորդականացվի, ինչը հնարավորություն կտա հետագա ուժեղացնել դրանք PCR-ի միջոցով և օգտագործել որպես զոնդ: STS-ի օգտագործումը հնարավոր կդարձնի դրանց հաջորդականությունները PCR արտադրանքների տեսքով օգտագործել որպես զոնդ՝ որոշակի գենոմային շրջանի ԴՆԹ-ի ցանկացած հատվածի նպատակային մեկուսացման համար գենոմային հաջորդականությունների գրադարանից: Արդյունքում կարող են ստեղծվել տվյալների բազաներ, որոնք ներառում են բոլոր STS-ների գտնվելու վայրը և կառուցվածքը, ինչպես նաև դրանց ուժեղացման համար անհրաժեշտ պրայմերները: Սա կվերացնի լաբորատորիաների՝ բազմաթիվ կլոններ պահելու և դրանք հետազոտության համար այլ լաբորատորիաներ ուղարկելու անհրաժեշտությունը: Բացի այդ, STS-ը հիմք է ստեղծում ընդհանուր լեզվի մշակման համար, որով տարբեր լաբորատորիաներ կարող են նկարագրել իրենց կլոնները: Այսպիսով, STS հայեցակարգի մշակման վերջնական արդյունքը կլինի մարդու գենոմի համապարփակ STS քարտեզը: Տեսականորեն 1 սմ չափի գենետիկական քարտեզ կառուցելու համար անհրաժեշտ է 3000 լիովին տեղեկատվական, պոլիմորֆ ԴՆԹ մարկերներ։ Այնուամենայնիվ, քանի որ պոլիմորֆ մարկերները անհավասարաչափ են բաշխված գենոմում և դրանցից միայն մի քանիսն են լիարժեք տեղեկատվական, այս չափի քարտեզ կառուցելու համար պահանջվող մարկերների իրական թիվը գնահատվում է 30-50 հազար: Ուսումնասիրվող քրոմոսոմային շրջաններին համապատասխանող մարկերներ ձեռք բերելու համար այժմ հաճախ օգտագործվում են ցրված կրկնվող հաջորդականություններին համապատասխան պրայմերներ, որոնց թվում առաջինը օգտագործվել են ալյուի հաջորդականությունները:
Alu-PCR.Ցրված կրկնվող ալուի հաջորդականությունները բնորոշ են մարդու գենոմին։ Alu-ի հաջորդականությունների համար հատուկ այբբենարաններ օգտագործվում են մարդու գենոմի ԴՆԹ-ի շրջանները, որոնք գտնվում են Alu-ի կրկնությունների միջև, որոնք գտնվում են միջինը 4-10 կբ հեռավորության վրա: միմյանցից. Alu-PCR-ի մեկ այլ տարբերակ է ԴՆԹ-ի զոնդերի նպատակային սինթեզը քրոմոսոմային շրջաններին, որոնք ստացվել են լազերային մասնատումից հետո, առանձին քրոմոսոմներին, որոնք մեկուսացված են հոսքի ցիտոմետրիայի միջոցով կամ հիբրիդային բջիջների ԴՆԹ-ին, որոնք պարունակում են մարդու գենոմի որոշակի մաս: Բացի այդ, Alu-PCR-ն օգտագործվում է բջջային հիբրիդներին գենոմի կայունության առումով բնութագրող եզակի մատնահետքեր ստանալու համար, ինչպես նաև բնութագրելու մարդու ԴՆԹ-ի բեկորները, որոնք կլոնավորված են YAC վեկտորներում, կոսմիդներում կամ բակտերիոֆագի ԴՆԹ-ի վրա հիմնված վեկտորներում: Մարդու գենոմի համար Ալուի հաջորդականությունների յուրահատկությունը հնարավորություն է տալիս դրանք օգտագործել «քրոմոսոմների երկայնքով քայլելու», ինչպես նաև գոյություն ունեցող կոնտիգների ընդլայնման համար: Քանի որ մարդկային գենոմում չափավոր կրկնվող հաջորդականությունների >90%-ը ներկայացված է Alu և KpnI ընտանիքներով, զարմանալի չէ, որ վերջիններս նույնպես օգտագործվում են PCR-ում նույն նպատակներով, ինչ Alu-ն: Այնուամենայնիվ, այստեղ PCR արտադրանքի պրոֆիլներն ավելի քիչ բարդ են, քանի որ KpnI հաջորդականությունները գենոմում ավելի քիչ են կրկնվում և ունեն բնորոշ տեղայնացում քրոմոսոմներում:
PCR-ն ակտիվորեն օգտագործվում է պոլիմորֆ մոլեկուլային մարկերների հայտնաբերման համար գենետիկ կապի քարտեզների կառուցման ժամանակ, որոնց հիմնական սկզբունքները քննարկվեցին վերևում: Այս մեթոդը նաև օգտակար է ԴՆԹ-ի հաջորդականացման և մարդու գենոմի բարձր լուծաչափով ֆիզիկական քարտեզների կառուցման գործում: PCR-ի կիրառման վերջին երկու ոլորտները ավելի մանրամասն կքննարկվեն ստորև:
Ցածր լուծման ֆիզիկական քարտեզներ
Ի տարբերություն վերը քննարկված գենետիկական կապի քարտեզների, գենոմի ֆիզիկական քարտեզներն արտացոլում են մարկերների միջև իրական հեռավորությունը՝ արտահայտված բազային զույգերով: Ֆիզիկական քարտեզները տարբերվում են իրենց լուծման աստիճանից, այսինքն. ըստ գենոմի կառուցվածքի մանրամասների, որոնք ներկայացված են դրանց վրա։ Առավելագույն լուծաչափով մարդու գենոմի համապարփակ ֆիզիկական քարտեզը կպարունակի նրա բոլոր քրոմոսոմների ամբողջական նուկլեոտիդային հաջորդականությունը: Նվազագույն լուծաչափով ֆիզիկական քարտեզների մյուս ծայրում են գենոմի քրոմոսոմային (ցիտոգենետիկ) քարտեզները:
Գենոմային ԴՆԹ-ի գենետիկ քարտեզների չորս տեսակներ և դրանց փոխհարաբերությունները

1 – գենետիկ կապի քարտեզ, 2 – ֆիզիկական սահմանափակման քարտեզ, բացատները ցույց են տալիս սահմանափակող ֆերմենտների միջոցով ԴՆԹ-ի տրոհման վայրերը, 3 – կոնտիգների ֆիզիկական քարտեզ, որը ցույց է տալիս YAC վեկտորների միջոցով ձեռք բերված համընկնող ԴՆԹ կլոնները, 4 – համապարփակ ֆիզիկական քարտեզ ԴՆԹ նուկլեոտիդային հաջորդականությունների տեսքով . Բոլոր քարտեզները ցույց են տալիս նույն քրոմոսոմային շրջանը
Քրոմոսոմային քարտեզներ.Մարդու գենոմի քրոմոսոմային քարտեզները ստացվում են առանձին քրոմոսոմների վրա գենետիկական մարկերների տեղայնացման միջոցով՝ օգտագործելով ցիտոգենետիկ մեթոդները, այդ թվում՝ ավտոռադիոգրաֆիա և FISH: Վերջին երկու դեպքերում ռադիոակտիվ կամ լյումինեսցենտային պիտակները, որոնք կապված են անձեռնմխելի քրոմոսոմների ուսումնասիրված գենետիկական վայրերի հետ, հայտնաբերվում են լուսային մանրադիտակի միջոցով: Վերջերս քրոմոսոմային քարտեզները հնարավորություն տվեցին տեղայնացնել ուսումնասիրվող ԴՆԹ-ի հատվածը 10 Մբ երկարությամբ քրոմոսոմային հատվածում։ Մետաֆազային քրոմոսոմների օգտագործմամբ in situ հիբրիդացման ժամանակակից մեթոդները, հիմնականում FISH մեթոդը, տեղայնացնում են պոլինուկլեոտիդային մարկերները 2-5 Մբ-ի սահմաններում: Ավելին, in situ հիբրիդացման ժամանակ ինտերֆազային քրոմոսոմների հետ, որոնցում գենետիկական նյութը պակաս կոմպակտ վիճակում է, քրոմոսոմային քարտեզների լուծունակությունը մոտենում է 100 կբ-ին։
Քրոմոսոմային քարտեզների ճշգրտությունը մեծանում է նաև ժամանակակից գենետիկական մեթոդների կիրառմամբ։ Օրինակ, մեկ սերմի ԴՆԹ հատվածները ուժեղացնելու PCR-ի կարողությունը թույլ է տալիս ուսումնասիրել մեծ թվով մեյոզներ, որոնք ասես պահպանված են առանձին սերմնահեղուկների նմուշներում: Արդյունքում հնարավոր է դառնում ստուգել քրոմոսոմային քարտեզների վրա տեղայնացված գենետիկական մարկերների հարաբերական դիրքը՝ օգտագործելով ավելի կոպիտ մեթոդներ։
cDNA քարտեզներ. cDNA քարտեզները արտացոլում են արտահայտված ԴՆԹ շրջանների (էկզոնների) դիրքը մետաֆազային քրոմոսոմների վրա հայտնի ցիտոգենետիկ մարկերների (զանգերի) նկատմամբ: Քանի որ նման քարտեզները պատկերացում են տալիս գենոմի տառադարձված շրջանների, ներառյալ անհայտ գործառույթներով գեների տեղայնացման մասին, դրանք կարող են օգտագործվել նոր գեներ որոնելու համար: Այս մոտեցումը հատկապես օգտակար է գեների որոնման ժամանակ, որոնց վնասը հանգեցնում է մարդու հիվանդությունների, եթե նման քրոմոսոմային շրջանների մոտավոր տեղայնացումը նախկինում արդեն իրականացվել է գենետիկ կապի քարտեզների վրա՝ ընտանեկան գենետիկական վերլուծության արդյունքում:
Բարձր լուծման ֆիզիկական քարտեզներ
ԴՆԹ-ի ֆիզիկական քարտեզներ կառուցելու երկու ռազմավարություն
ա – «վերևից ներքև» ռազմավարություն. ամբողջ քրոմոսոմի ԴՆԹ-ն ճեղքվում է մեծ կտրվածքով սահմանափակող ֆերմենտների միջոցով և կառուցվում է սահմանափակումային քարտեզ յուրաքանչյուր առանձին ԴՆԹ-ի բեկորների համար. բ – ներքևից վեր ռազմավարություն, YAC-ի անհատական կլոնները նույնականացումից հետո համակցվում են կոնտիգների
Մարդու գենոմի բարձր լուծաչափով քարտեզներ ստեղծելու փորձերում փորձարարականորեն իրականացվում են երկու այլընտրանքային մոտեցումներ, որոնք կոչվում են վերից վար և ներքևից վեր քարտեզագրում: Վերևից ներքև քարտեզագրելիս վերլուծության մեկնարկային կետը մարդու անհատական քրոմոսոմի ԴՆԹ-ի նմուշն է: ԴՆԹ-ն կտրվում է մեծ կտրվածքով սահմանափակող ֆերմենտների միջոցով (օրինակ՝ NotI) երկար բեկորների, որոնք իմպուլսային էլեկտրական դաշտում էլեկտրոֆորեզով բաժանվելուց հետո ենթարկվում են սահմանափակման հետագա վերլուծության այլ սահմանափակող ֆերմենտների հետ: Արդյունքում ստացվում է մակրոսահմանափակման քարտեզ, որը բավականին լիովին ներկայացնում է ուսումնասիրվող քրոմոսոմի կամ նրա մասերի բոլոր հաջորդականությունները, սակայն դրա թույլտվությունը ցածր է։ Նման քարտեզի վրա շատ դժվար է տեղայնացնել առանձին գեներ։ Բացի այդ, յուրաքանչյուր առանձին քարտեզ հազվադեպ է ընդգրկում ԴՆԹ-ի ընդլայնված հատվածները (սովորաբար ոչ ավելի, քան 1-10 Մբ):
Մարդու գենոմը ներքևից վերև քարտեզագրելիս՝ հիմնվելով գենոմի կամ առանձին քրոմոսոմի ընդհանուր ԴՆԹ-ի պատրաստման վրա, ընդլայնված ԴՆԹ հաջորդականությունների մի շարք պատահական կլոններ (10–1000 կբ), որոնցից մի քանիսը համընկնում են միմյանց հետ, ստացվում են. Բակտերիալ արհեստական մինիքրոմոսոմները (BAC) կամ խմորիչ արհեստական քրոմոսոմները (YAC), որոնք մանրամասն նկարագրված են 7.2.4 բաժնում, հաճախ օգտագործվում են որպես կլոնավորման վեկտոր այս դեպքում: Մասամբ համընկնող և փոխլրացնող կլոնների շարքը կազմում է ԴՆԹ-ի նուկլեոտիդների շարունակական հաջորդական հաջորդականություն, որը կոչվում է կոնտիգ։ Ստացված կոնտիգների ճիշտությունը հաստատվում է in situ հիբրիդացումով (FISH)՝ ուսումնասիրվող քրոմոսոմների որոշակի շրջանների հետ դրանց միաժամանակյա կապով։ Contig-ի վրա հիմնված քարտեզները ամբողջական տեղեկատվություն են տալիս քրոմոսոմի առանձին հատվածների կառուցվածքի մասին և թույլ են տալիս տեղայնացնել առանձին գեները: Այնուամենայնիվ, նման քարտեզները դժվար է օգտագործել ամբողջ քրոմոսոմների կամ դրանց ընդլայնված հատվածների վերակառուցման համար՝ հասանելի գեների գրադարանում համապատասխան կլոնների բացակայության պատճառով։
Հիմնական խնդիրը, որը պետք է լուծվի բարձր լուծաչափով ֆիզիկական քարտեզների կառուցման երկու մոտեցումներն էլ կիրառելիս, ցրված ԴՆԹ-ի բեկորների համակցումն է շարունակական նուկլեոտիդային հաջորդականությունների մեջ: Ամենից հաճախ դրա համար օգտագործվում են հատուկ կլոնավորված ԴՆԹ բեկորներ, որոնք կոչվում են կապող կլոններ: Կապակցող կլոններից ԴՆԹ-ի բեկորները իրենց ներքին մասերում պարունակում են մեծ կտրվածքով սահմանափակող ֆերմենտների նուկլեոտիդային հաջորդականություններ և, հետևաբար, ներկայացնում են ֆիզիկական քարտեզագրման առաջին փուլերում օգտագործվող ԴՆԹ-ի բեկորների միացման վայրերը: Հարավային հիբրիդացումը, որն օգտագործում է միացնող կլոնների ԴՆԹ-ի բեկորները որպես զոնդ, որոշում է ֆիզիկական քարտեզների ԴՆԹ-ի բեկորները, որոնք պարունակում են նուկլեոտիդային հաջորդականություններ մեծ կտրվածքով սահմանափակող ֆերմենտների սահմանափակման վայրերի մոտակայքում: Եթե հայտնաբերվում են երկու նման բեկորներ, ապա համապատասխան կապող կլոնը համընկնում է և մտնում է այս երկու բեկորների մեջ։ Իր հերթին, կապող կլոնները ընտրվում են գենային գրադարաններից՝ օգտագործելով զոնդերը, որոնք մեծ կտրվածքով սահմանափակող ֆերմենտների սահմանափակման վայրերի նուկլեոտիդային հաջորդականություններ են:
ՑԱՆԿ ՕԳՏԱԳՈՐԾՎԱԾ ԱՂԲՅՈՒՐՆԵՐ
1) Քլարկ Մ.Ս. Համեմատական գենոմիկա. Մարդու գենոմի նախագիծը հասկանալու բանալին // BioEssays. 1999. Հատ. 21. Էջ 21–30։
2) Billings P.R., Smith C.L., Cantor C.L. Մարդու գենոմի ֆիզիկական քարտեզագրման նոր տեխնիկա // FASEB J. 1991. Vol. 5. Էջ 28–34։
3) Գեորգիև Գ.Պ. Բարձրագույն օրգանիզմների գեները և դրանց արտահայտությունը. M.: Nauka, 1989. 254 p.
4) http://referatwork.ru/refs/source/ref-8543.html
Բժշկության հետ կապված մոլեկուլային գենետիկայի ամենակարևոր խնդիրը մարդու ժառանգական հիվանդությունների գեների նույնականացումն է և դրանցում հատուկ վնասների հայտնաբերումը, ինչը հանգեցնում է հիվանդության ֆենոտիպային դրսևորումների զարգացմանը: Այս առաջադրանքը կարող է իրականացվել մի քանի հիմնական ենթակետերի միջոցով.
\ՕԴՈՎ.
Գենի նույնականացման առաջին մոտեցումը, որը առաջատար մնաց մինչև 90-ականների սկիզբը,
| հետազոտվող հիվանդությունը բնութագրող հիմնական կենսաբանական թերության (գենի առաջնային սպիտակուցային արտադրանքի) մասին առկա տեղեկատվության հիման վրա | Շիշկին Ս.Ս., Կալինին Վ.Ն., ] 992; Gardner E. et al., 1991; Collins F., 1995]:
Սպիտակուցի վերլուծությունից ԴՆԹ մակարդակի անցումը կատարվել է մաքրված սպիտակուցային արտադրանքի հաջորդականացման և ԴՆԹ-ի զոնդերի ստացման, մոնոկլոնալ հակամարմինների օգտագործման և որոշ այլ մեթոդաբանական տեխնիկայի միջոցով: Գենի քրոմոսոմային տեղայնացումը այս որոնման մեջ սխեման հետազոտության վերջնական արդյունքն է:Նկարագրված մոտեցումը, օգտագործելով ցանկալի գենի ֆունկցիոնալ նշանակության այս կամ այն նախնական տեղեկությունը, կոչվում է «ֆունկցիոնալ կլոնավորում»:Ֆունկցիոնալ կլոնավորման հաջող կիրառման օրինակ է ֆենիլկետոնուրիայի գենի նույնականացումը: Ցավոք, այս մեթոդը կարող է կիրառվել միայն մարդկային հիվանդությունների շատ սահմանափակ շրջանակի համար, մինչդեռ ժառանգական հիվանդությունների մեծ մասի համար առաջնային գենային արտադրանքները կամ պաթոգոմոնիկ կենսաքիմիական մարկերները անհայտ են:
Մոլեկուլային տեխնոլոգիաների բարելավումները հանգեցրել են գեների որոնման հիմնովին այլ ռազմավարության ստեղծմանը, որը չի պահանջում որևէ նախնական իմացություն իր ֆունկցիայի կամ առաջնային կենսաքիմիական արտադրանքի մասին: Այս ռազմավարությունը ենթադրում է գենի նույնականացում՝ հիմնված կոնկրետ քրոմոսոմային տեղայնացման ճշգրիտ իմացության վրա՝ «դիրքային կլոնավորում» (ավելի քիչ հաջողված «հակադարձ գենետիկա» տերմինը): Դիրքային կլոնավորումը հանգեցնում է հիվանդության մոլեկուլային հիմքի հաստատմանը «գենից մինչև սպիտակուց» և ներառում է հետևյալ հիմնական փուլերը. 2) ուսումնասիրված քրոմոսոմային շրջանի ֆիզիկական քարտեզի կազմում (ֆիզիկական քարտեզագրում). 3) ուսումնասիրված տարածաշրջանում արտահայտված ԴՆԹ-ի հաջորդականությունների նույնականացում. 4) թեկնածու գեների հաջորդականություն և հիվանդ անհատների մոտ ցանկալի գենի մուտացիաների հայտնաբերում. 5) գենային կառուցվածքի վերլուծություն.
վերծանելով իր արտադրանքի հաջորդականությունը և առաջնային կառուցվածքը՝ mRNA և սպիտակուց: Որոշ դեպքերում գենի դիրքային կլոնավորումը հեշտանում է, երբ հիվանդների մոտ հայտնաբերվում են տեսանելի ցիգոգենետիկ վերադասավորումներ կամ հայտնաբերվող ջնջումներ կրիտիկական քրոմոսոմային շրջանում, ինչը կարող է զգալիորեն մեծացնել մուտանտի գենի քարտեզագրման ճշգրտությունը: Նման վերադասավորումների բացահայտումը, մասնավորապես, նպաստեց Դյուշենի/Բեկերի մկանային դիստրոֆիայի, 1-ին տիպի նեյրոֆիբրոմատոզի, տուբերոզ սկլերոզի, ադրենոլեյկոդիստրոֆիայի և նյարդային համակարգի այլ ժառանգական հիվանդությունների գեների կլոնավորման հաջողությանը:
«Մարդու գենոմ» հետազոտական նախագծի կարևոր միջանկյալ արդյունքներից մեկը գենոմի ավելի ու ավելի հարուստ տրանսկրիպցիոն քարտեզի ստեղծումն էր, որը պարունակում է տեղեկատվություն հազարավոր արդեն հայտնի գեների և արտահայտված նուկլեոտիդային հաջորդականությունների մասին: Սա նպաստել է առաջնային գենետիկական արատը բացահայտելու մեկ այլ մոտեցման էական զարգացմանը, որում մուտանտ գենի նախնական քարտեզագրումից հետո ստուգվում են նույն քրոմոսոմային շրջանում տեղակայված համապատասխան թեկնածու գեները («դիրքային թեկնածուի մոտեցում»): Այս մեթոդը ենթադրում է որոշակի գիտելիքներ ուսումնասիրվող հիվանդության պաթոֆիզիոլոգիայի մասին, ինչը հնարավորություն է տալիս ռացիոնալ կերպով ընտրել վերլուծության համար թեկնածու գեները մեծ թվով գեներից, որոնք կարող են տեղակայվել «հետաքրքրության գոտում»: Նյարդաբանական ժառանգական հիվանդությունների շարքում, որոնց գեները այս կերպ բացահայտվել են համապատասխան թեկնածուների վերլուծության միջոցով սահմանված քրոմոսոմային միջակայքում, դոպա-կախյալ դիստոնիան և Ֆրիդրեյխի նման ատաքսիան վիտամին E-ի պակասով: Ըստ ընթացիկ կանխատեսումների, դա վերլուծություն է: դիրքային թեկնածուներ», որը մոտ ապագայում կդառնա զարգացման կիզակետը ժառանգական հիվանդությունների գեների նույնականացման առաջատար մեթոդ, որին մեծապես նպաստում է արտահայտված հաջորդականության պիտակների համակարգչային տվյալների բազաների ստեղծումը և մշտական ընդլայնումը:
Այսպիսով, ցանկալի գենի քրոմոսոմային տեղայնացման որոշումը՝ գենետիկական քարտեզագրումը, առաջին, առանցքային քայլն է որոշակի ժառանգական հիվանդության մոլեկուլային հիմքի բացահայտման ուղղությամբ:
Կան մի քանի հիմնական մեթոդներ, որոնք թույլ են տալիս քարտեզագրել անհայտ գենը կոնկրետ քրոմոսոմային տեղանքում. Օրինակ՝ X քրոմոսոմի վրա գենի տեղայնացման հաստատումը X-կապակցված տիպի միջոցով հիվանդության փոխանցման դեպքում. բ) ցիտոգենետիկ - հիմնված է մանրադիտակի միջոցով հայտնաբերված քրոմոսոմային վերադասավորումների միացման վրա որոշակի կլինիկական ֆենոտիպի հետ. գ) in situ հիբրիդացման մեթոդը (ներառյալ դրա ժամանակակից մոդիֆիկացիան՝ լյումինեսցենտ in situ հիբրիդացում, FISH) - օգտագործում է ցանկալի գենի mRNA-ի և cDNA-ի հատուկ հիբրիդացում՝ դենատուրացված քրոմոսոմներով մետաֆազային բջիջների պատրաստուկների վրա. դ) հիբրիդային բջիջների մեթոդ - հիմնված է in vitro կլոնավորված հիբրիդային սոմատիկ բջիջներում բջջային բնութագրերի և քրոմոսոմների համատեղ տարանջատման վերլուծության վրա [Fogel F., Motulski A., 1990; Gardner E. et al., 1991]: Այս բոլոր մեթոդները գտել են իրենց կիրառությունը ժամանակակից մոլեկուլային գենետիկայի մեջ, սակայն դրանք ունեն լուրջ սահմանափակումներ՝ կապված ինչպես անբավարար լուծման, այնպես էլ հետազոտություններ անցկացնելու համար անհրաժեշտ խիստ նախադրյալների առկայության հետ (օրինակ՝ զոնդերի առկայությունը, հիբրիդային բջիջների ընտրության ընտրովի համակարգերի առկայությունը։ և այլն): Մարդու ժառանգական հիվանդությունների գեների քարտեզագրման ամենահզոր, արդյունավետ և ներկայումս լայնորեն օգտագործվող մեթոդը այսպես կոչված կապի վերլուծությունն է՝ ցանկալի գենի կապի վերլուծությունը ճշգրիտ տեղայնացված գենետիկական մարկերների մի շարքով:
Կապի վերլուծության կենտրոնական դիրքն այն է, որ քրոմոսոմի վրա երկու տեղամասերի միջև հարաբերական գենետիկական հեռավորության չափումը կարող է լինել այս տեղանքների միջև վերահամակցումների հաճախականությունը մեյոզում հոմոլոգ քրոմոսոմների հատման արդյունքում: Որքան մոտ են տեղանքները տեղակայված քրոմոսոմի վրա, այնքան մեծ է հավանականությունը, որ դրանք կժառանգվեն որպես մեկ ամբողջություն (կապող խումբ); եթե ուսումնասիրված տեղանքները զգալիորեն հեռու են (այսինքն՝ ունեն կապի թույլ աստիճան), ապա դրանք ավելի հավանական է, որ ցրվեն տարբեր քրոմոսոմներ անցնելուց հետո: Լոկերի միջև ռեկոմբինացիայի հաճախականությունը 1% ընդունվում է որպես միասնություն
Բրինձ. 30. Աուտոսոմային գերիշխող հիվանդության օրինակով գենետիկ կապի վերլուծության սկզբունքը Այս օրինակում ուսումնասիրվել են A, B, C և D 4 կապակցված մարկերներ, որոնցից վերակառուցվել են հապլոտիպերը: Տարբեր ծագման քրոմոսոմները նշվում են տարբեր տեսակի ստվերներով (բնօրինակ մուտանտի քրոմոսոմը նշված է սևով): Տոհմային բոլոր հիվանդներն ունեն սկզբնական մուտանտի քրոմոսոմի նույն ընդհանուր (միջին) մասը: Օրինակ, ստորին սերնդում քրոմոսոմները ենթարկվել են մի շարք վերակոմբինացիաների, բայց բոլոր հիվանդ սիբները (ներառյալ անհատները Sh-Z և Sh-8) պահպանում են նույն մուտանտի հապլոտիպը B և C մարկերների համար (haplotype y): Ընդհակառակը, ցածր սերնդի առողջ քույրերից և եղբայրներից և ոչ մեկը չի ժառանգել հապլոտիպ j իրենց հորից B և C մարկերների համար (անհատական Ш-4-ը ժառանգել է քրոմոսոմ, որտեղ վերահամակցումը տեղի է ունեցել կրիտիկական հատվածից ցածր): Այսպիսով, մարկերային ալելների տարանջատումը և հապլոտիպերի վերլուծությունը ցույց են տալիս, որ հիվանդության գենը գտնվում է քրոմոսոմային հատվածում, որը ներառում է B և C մարկերներ: Համապատասխանաբար, քրոմոսոմային շրջանի արտաքին սահմանները, որոնցում գտնվում է մուտանտի գենը, մարկերներն են A և D:
և ուսումնասիրվող մարկերի նույն ալելը, սա ցույց է տալիս ցանկալի մուտանտի գենի և այս մարկերի միջև ռեկոմբինացիաների բացակայությունը, այսինքն. նրանց միջև կպչունության առկայության մասին. Աուտոսոմային գերիշխող հիվանդության գենի և որոշակի գենետիկ մարկերների միջև կապի օրինակ ներկայացված է Նկ. երեսուն.
Համախմբվածությունը հուսալիորեն ապացուցելու համար մշակվել է հատուկ մաթեմատիկական ապարատ։ Հաշվարկի սկզբունքն է համեմատել կապի առկայության և բացակայության մասին վարկածների հավանականությունը ընտանիքի առկա տվյալների և ընտրված ռեկոմբինացիայի հաճախականության հետ 0; Այս երկու հավանականությունների հարաբերակցությունը (հավանականության հարաբերակցություն) արտահայտում է կապի կողմ և դեմ հավանականությունը: Հարմարության համար օգտագործվում է հավանականության հարաբերակցության տասնորդական լոգարիթմը՝ հավանականության լոգարիթմը կամ LOD.
Po
LOD = Logio --
P1/2, որտեղ P-ը հավանականություն է
0-ի ռեկոմբինացիայի հաճախականությամբ կապակցված գեների ընտանիքի տվյալների բաշխումը, P-ն այդպիսի բաշխման հավանականությունն է երկու չկապված ազատորեն վերամիավորվող գեների համար (վերակոմբինացիայի հաճախականությունը 0 = 1/2): Հաշվարկի լոգարիթմական ձևի օգտագործումը թույլ է տալիս ավելացնել առանձին տոհմերի վերլուծությունից ստացված 27od կետերը: Գենետիկական կապն ապացուցելու համար ընդունվել է +3 Lod միավորը, ինչը նշանակում է 1000:1 գործակից՝ հօգուտ gt-ի միջև գենետիկ կապի առկայության; մարկեր և ուսումնասիրվող բնութագիր: Lod միավորը -2 և ցածր ցույց է տալիս զուգավորման զգալի բացակայություն; Կպչունության առկայության տեսանկյունից Lod-score արժեքները +3-ից մինչև -2, համապատասխանաբար, քիչ թե շատ ենթադրյալ են և պահանջում են հետագա հաստատում: Ռեկոմբինացիայի հաճախականությունը 0, որի համար սահմանվել է առավելագույն L od միավորը, ուսումնասիրված տեղանքների միջև ամենահավանական գենետիկական հեռավորության արտացոլումն է. Մոտավորապես ենթադրվում է, որ ռեկոմբինացիաների 1%-ը ցույց է տալիս շատ սերտ կապ, մոտ 5% ռեկոմբինացիայի հաճախականությունը ցույց է տալիս լավ կապ, իսկ 10-20% հաճախականությունը ցույց է տալիս որոշ չափավոր կապ:
Lob կետերի հաշվարկը ներառում է հատուկ համակարգչային ծրագրերի օգտագործում (LIPED ծրագիր, LINKAGE ծրագրային փաթեթ և այլն):
Որպեսզի կապի վերլուծությունը հաջող լինի, անհրաժեշտ է, որ ուսումնասիրվող ընտանիքները տեղեկացված լինեն հիվանդության և գենետիկական մարկերի մասին: Առաջինը նշանակում է տոհմում բավարար քանակությամբ տեղեկատվական մեյոզների առկայություն, ինչը թույլ է տալիս վերլուծել տվյալ տոհմային նիշերի տարբերությունը: Գործնական տեսանկյունից սա նշանակում է ունենալ մեծ թվով հիվանդ և առողջ հարազատներ, որոնք հասանելի են վերլուծության համար, որոնք սովորաբար ընդգրկում են մի քանի սերունդ: Մարկերի տեղեկատվական լինելը ենթադրում է նրա պոլիմորֆիզմը (այսինքն՝ մեծ թվով ալելների առկայություն) և հետերոզիգոտություն ընտանիքի հիմնական անդամների մոտ, ինչը հնարավորություն է տալիս տարբերակել հատուկ մարկերային ալելների գենետիկական ծագումը։ Մինչև 1980-ականների վերջը կապի վերլուծության մեջ օգտագործվող մարկերների հիմնական տեսակը քրոմոսոմային ԴՆԹ-ի հատվածներն էին, որոնք պարունակում էին տատանումներ մեկ բազային զույգում և տարբերվում էին համապատասխան ֆերմենտի սահմանափակման վայրի առկայությամբ կամ բացակայությամբ, այսինքն. սահմանափակող բեկորների երկարությամբ («սահմանափակող հատվածի երկարության պոլիմորֆիզմ», RFLP): Գենետիկ քարտեզագրման նոր դարաշրջանը սկսվել է բարձր պոլիմորֆ մարկերների դասի հայտնաբերմամբ, որոնք ԴՆԹ-ի հատվածներ են, որոնք բաղկացած են տանդեմ (SA)n կրկնվող կրկնօրինակների փոփոխական թվով և ունեն չափազանց բարձր հետերոզիգոտություն: Սա հնարավորություն տվեց մեծապես լուծել օգտագործված մարկերների տեղեկատվական բովանդակության խնդիրը և նպաստեց կապի վերլուծության զգալի առաջընթացին: Ըստ որոշ գնահատականների, ամբողջական հապլոիդ գենոմը զննելու և գենետիկական կապը բացահայտելու համար անհրաժեշտ է ունենալ 200-300 բարձր պոլիմորֆ մարկերներ, որոնք հավասարապես բաշխված են քրոմոսոմների երկայնքով: Վերջին սերնդի գենետիկական քարտեզները ներառում են ավելի քան 5000 այդպիսի մարկերներ, ինչը թույլ է տալիս մեզ այսօր դիտարկել գենետիկական կապի հաստատման խնդիրը, որը հիմնովին հնարավոր է ցանկացած տեղեկատվական ծագումնաբանության մեջ:
Լուրջ խնդիր, որը հանդիպում է մի շարք ընտանիքների վրա կապի վերլուծություն իրականացնելիս, ուսումնասիրվող կլինիկական համախտանիշի հնարավոր գենետիկական տարասեռության խնդիրն է: Եթե ուսումնասիրվող ֆենոտիպը կարող է առաջանալ տարբեր գեների մուտացիաներով, ապա առանձին ընտանիքներում ստացված դրական (կապակցման առկայության դեպքում) և բացասական (կապման բացակայության դեպքում) Lod միավորների մեխանիկական ավելացումը հանգեցնում է ընդհանուր Lod-ի համահարթեցմանը: միավորի արժեք և կեղծ եզրակացություն կապի իսպառ բացակայության մասին: Օրինակ՝ 1-ին տիպի ավտոսոմալ գերիշխող շարժիչ-զգայական նյարդաբանությունը, որն առաջանում է 1-ին, 17-րդ և այլ քրոմոսոմներում տեղայնացված տարբեր գեների մուտացիաների հետևանքով: Այս իրավիճակում կապի վերլուծության համար ուղարկված հիվանդների և ընտանիքների մանրակրկիտ, մանրակրկիտ հետազոտությունն առանձնահատուկ նշանակություն ունի՝ առավել միատարր կլինիկական խմբերը ընտրելու համար: Կեղծ բացասական թեստի արդյունքից խուսափելու լրացուցիչ միջոց օգտագործելն է
ta,/7od-կետերը հատուկ HOMOG ծրագրի կամ նմանատիպ ծրագրերի, որոնք թույլ են տալիս գնահատել գենետիկական տարասեռության հավանականությունը ընտանեկան տվյալների որոշակի փաթեթի համար: Ուսումնասիրության առաջին փուլում ամենաարդյունավետ մոտեցումը կապի վերլուծությունն է մեկ մեծ տեղեկատվական տոհմում, ինչը թույլ է տալիս դիտավորյալ բացառել հիվանդների ուսումնասիրված խմբի գենետիկական տարասեռության հնարավորությունը: Կապի վերլուծության ժամանակ լրացուցիչ դժվարությունները կապված են մուտանտի գենի հաճախ նկատվող թերի ներթափանցման և փոփոխական արտահայտչականության, հետազոտված ընտանիքի անդամների միջև ֆենոկոպիայի առկայության, հիվանդության առաջացման տարիքի գնահատման և մուտացիայի նախակլինիկական փոխանցման հնարավորության հետ: , պոպուլյացիայի մեջ ուսումնասիրվող մարկերների հատուկ ալելների տարածվածության գնահատում և այլն։ . Այս գործոնների սխալ հաշվառումը կամ թերագնահատումը կարող է էապես ազդել վերջնական արդյունքի վրա, հետևաբար առաջին պլան է մղվում ուսումնասիրվող ընտանիքներում մանրամասն կլինիկական և ծագումնաբանական վերլուծության որակը:
Մշակվել են բազմաթիվ նոր մեթոդներ, որոնք ներկայացնում են գենետիկ կապի ուսումնասիրման ավանդական ռազմավարության հետագա զարգացումը և զգալիորեն մեծացնում են կատարման արագությունը, մեթոդաբանական հնարավորությունները և այս վերլուծության լուծումը ժառանգական մարդու հիվանդությունների անհայտ գեների տեղայնացման հարցում: Այս մեթոդներից մեկը բազմաբնույթ կապի վերլուծությունն է, որը թույլ է տալիս գնահատել Lod-ի միավորները մի շարք կապակցված տեղանքների համար՝ համաձայն ուսումնասիրվող քրոմոսոմային շրջանի գենետիկական քարտեզի և որոշել մուտանտի գենի ամենահավանական տեղայնացումը այս տարածաշրջանում: Ինբրեդում
Աուտոսոմային ռեցեսիվ հիվանդությամբ տոհմերը «հիմնադիր էֆեկտի» ենթադրության առկայության դեպքում, հոմոզիգոտ քարտեզագրման մեթոդն իրեն չափազանց արդյունավետ է ցույց տվել. ախտահարված անհատների կողմից մի շարք մարկերներ՝ որպես ընդհանուր քրոմոսոմային շրջանի մեկ նախնիից ժառանգության արդյունքում, որը ներառում է մուտանտ գեն: «Տնտեսական գենոմի սկանավորման» մեթոդը խոստումնալից է, որը ներառում է արտահայտված հաջորդականությամբ հարուստ «ռազմավարական» CG հագեցած քրոմոսոմային շրջաններում տեղայնացված մարկերների արտոնյալ օգտագործումը: Առաջարկվել են նաև դասական կապի վերլուծության մի շարք այլ փոփոխություններ:
Կարևոր է ընդգծել, որ կապի վերլուծությունը կարևոր կմնա նույնիսկ ամբողջ մարդկային գենոմի նույնականացումից հետո: Օրինակ, չբացահայտված գեներով ժառանգական հիվանդությունների դեռևս բավականին մեծ խումբ ուսումնասիրելիս, մոլեկուլային արատը պարզելու առաջին քայլը կարող է լինել գենետիկական վերլուծությունը և հիվանդության քրոմոսոմային տեղանքի նույնականացումը, որին հաջորդում է այս տարածաշրջանում համապատասխան գեների սքրինինգը: Բժշկական բժշկի դերը չափազանց կարևոր է գենետիկ քարտեզագրման հաջողության համար: Այն բաղկացած է ներկայացուցչական ընտանիքների համապատասխան ընտրությունից, հետազոտության մեջ ընդգրկված ընտանիքի բոլոր անդամների կլինիկական կարգավիճակի մանրամասն գնահատումից, հիվանդության ճշգրիտ ախտորոշումից և մուտանտի գենի տարանջատման օրինաչափության գնահատումից, ինչպես նաև բազմաթիվ այլ հիմնական խնդիրների լուծումից:
Մարդու գենոմի քարտեզագրում
Մենք կարիք չունենք իզուր անհանգստացնել աստվածներին,
Պատերազմի մասին գուշակելու համար զոհերի ներքին մտքեր կան,
Ստրուկներ՝ լռելու և քարեր՝ կառուցելու։
Օսիպ Մանդելշտամ, «Բնությունը նույնն է, ինչ Հռոմը…»
Գենետիկան երիտասարդ գիտություն է։ Տեսակների էվոլյուցիան իսկապես հայտնաբերվեց միայն 19-րդ դարի 50-ականների վերջին: 1866 թվականին ավստրիացի վանական Գրեգոր Մենդելը հրապարակեց սիսեռի փոշոտման վերաբերյալ իր փորձերի արդյունքները։ Մինչև դարի վերջը ոչ ոք ուշադրություն չդարձրեց նրա հայտնագործությանը։ Իսկ Գալթոնը, օրինակ, երբեք չգիտեր դրանց մասին։ Նույնիսկ բեղմնավորման մեխանիզմը՝ արական և իգական սեռական բջիջների միջուկների միաձուլումը, հայտնաբերվել է միայն 1875 թվականին։ 1888 թվականին բջիջների միջուկներում հայտնաբերվեցին քրոմոսոմներ կոչվող մարմիններ, իսկ 1909 թվականին Մենդելյան ժառանգական գործոնները կոչվեցին գեներ։ Առաջին արհեստական բեղմնավորումը (նապաստակի, ապա կապիկների մոտ) կատարվել է 1934թ. և վերջապես 1953 թվականին արվեց հիմնարար հայտնագործություն՝ հաստատվեց ԴՆԹ-ի կրկնակի պտուտակավոր կառուցվածքը։ Ինչպես տեսնում ենք, այս ամենը տեղի ունեցավ բոլորովին վերջերս, այնպես որ վաղ եվգենիկները, ընդհանուր առմամբ, շատ քիչ տեղեկացված էին իրենց աշխատանքի տեխնոլոգիայի մասին։
Մարդու գենոմի քարտեզագրումը դեռ վաղ փուլերում է։ Այն, ինչ մենք գիտենք, փոքր մասնաբաժին է՝ համեմատած այն, ինչ մենք չգիտենք: Կան երեք միլիարդ նուկլեոտիդային հաջորդականություններ, որոնք ձևավորվում են քսանվեցից մինչև երեսունութ հազար գեներ, որոնք ուղղակիորեն կոդավորում են սպիտակուցները: Բայց թե ինչպես են փոխազդում գեներն ու նրանց արտադրած սպիտակուցները, դեռևս վատ է հասկացվում:
Այնուամենայնիվ, գեների դերը մարդկային հասարակության մեջ բավականին արագ է գիտակցվում։ 1998 թվականին Դիանա Փոլը (Մասաչուսեթսի համալսարան) հիշեց, որ տասնչորս տարի առաջ նա զանգահարել էր.
«կենսաբանորեն դետերմինիստական» տեսակետ, որ ինտելեկտի և խառնվածքի տարբերությունների վրա ազդում են գեները՝ օգտագործելով այս տերմինները, կարծես թե դրանց նշանակությունը հստակեցված է: Այսօր դրանց օգտագործումը հակասական կլիներ, քանի որ այս պիտակները կարծես թե կասկածի տակ են դնում այս տեսակետը, մինչդեռ այն լայնորեն ընդունված է ինչպես գիտնականների, այնպես էլ հանրության կողմից»:.
Ինչ էլ որ լինի, մեր գիտելիքները բառացիորեն ամեն օր ընդլայնվում են, և շատ մոտ ապագայում մենք կկարողանանք մեծ ճշգրտությամբ վերլուծել. գենետիկական բեռորը մենք պարտադրում ենք ապագա սերունդներին։
Փաստերի նորագույն գիրքը գրքից: Հատոր 1 [Աստղագիտություն և աստղաֆիզիկա. Աշխարհագրություն և երկրային այլ գիտություններ։ Կենսաբանություն և բժշկություն] հեղինակ «Մարդկային գենոմը. չորս տառով գրված հանրագիտարան» գրքից հեղինակ Մարդու գենոմը գրքից [Հանրագիտարան գրված չորս տառով] հեղինակ Տարանտուլ Վյաչեսլավ Զալմանովիչ Փաստերի նորագույն գիրքը գրքից: Հատոր 1. Աստղագիտություն և աստղաֆիզիկա. Աշխարհագրություն և երկրային այլ գիտություններ։ Կենսաբանություն և բժշկություն հեղինակ Կոնդրաշով Անատոլի Պավլովիչ Life Deciphered [Իմ գենոմը, իմ կյանքը] գրքից Վենտեր Քրեյգի կողմից Կենսաբանական քիմիա գրքից հեղինակ Լելևիչ Վլադիմիր Վալերյանովիչ Հեղինակի գրքից Հեղինակի գրքիցՄԱՍ I. ՄԱՐԴՈՒ ԳԵՆՈՄԻ ԿԱՌՈՒՑՎԱԾՔԸ ԻՆՉ Է ԳԵՆՈՄԸ: Հարցերը հավերժ են, պատասխանները որոշվում են ժամանակով։ E. Chargaff Կյանքի հետ երկխոսության մեջ կարևոր է ոչ թե նրա հարցը, այլ մեր պատասխանը: M. I. Tsvetaeva Հենց սկզբից եկեք սահմանենք, թե ինչ ենք հասկանում այստեղ գենոմ բառով: Այս տերմինն ինքնին
Հեղինակի գրքիցԸնդհանուր ԴՆԹ-ի վերլուծություն՝ նոր տեղեկատվություն մարդու գենոմի կառուցվածքի մասին Մարդու գենոմի կառուցվածքի ուղղակի հետազոտության առաջին փուլում, երբ գենետիկական ինժեներիայի մեթոդաբանությունը դեռ գոյություն չուներ, ԴՆԹ-ի ուսումնասիրության համար կիրառվեցին ավանդական ֆիզիկաքիմիական մեթոդներ: IN
Հեղինակի գրքից Հեղինակի գրքիցՄԱՍ II. ՄԱՐԴՈՒ ԳԵՆՈՄԻ ՖՈՒՆԿՑԻԱ ԹԱԳՈՒՀԸ ՄԵՌԱՑՎԱԾ Է. ԿԵՑՑԻ ԹԱԳՈՒՀԸ: Այն, ինչ մենք գիտենք, սահմանափակ է, բայց այն, ինչ մենք չգիտենք, անսահման է: P. Laplace Գիտությունը միշտ էլ սխալ է ստացվում: Նա երբեք չի լուծի որևէ խնդիր՝ առանց տասնյակ նորերի բարձրաձայնելու: Բ. Շոու Այսպիսով,
Հեղինակի գրքիցԻնչպե՞ս է համակարգիչը օգտակար մարդու գենոմը ուսումնասիրելու համար: Առանց համակարգչային կենսատեղեկատվական տեխնոլոգիաների (գենոինֆորմատիկա կամ, ավելի լայն իմաստով, կենսաինֆորմատիկա) գենոմային հետազոտությունների զարգացումը դժվար թե ընդհանրապես հնարավոր լիներ։ Նույնիսկ դժվար է պատկերացնել, թե ինչպես
Հեղինակի գրքիցՄԱՍ III. ՄԱՐԴՈՒ ԳԵՆՈՄԻ ԾԱԳՈՒՄԸ ԵՎ ԷՎՈԼՈՒՑԻԱ
Հեղինակի գրքիցՈրքանո՞վ է տարբերվում մարդու գենոմը շիմպանզեի գենոմից: Գենոմը գեների հավաքածու է, որը պարունակվում է տվյալ օրգանիզմի քրոմոսոմների հապլոիդ (մեկ) հավաքածուում։ Գենոմը ոչ թե անհատի, այլ օրգանիզմի տեսակի հատկանիշ է։ 2001 թվականի փետրվարին ամերիկ
Հեղինակի գրքիցԳլուխ 11 Մարդու գենոմի վերծանում Ի՞նչ եք ասում, երբ ամբողջ ուժով բարձրանալով մի լեռան գագաթ, որտեղ ոչ ոք երբեք չի եղել, հանկարծ տեսնում եք, թե ինչպես է մարդ բարձրանում զուգահեռ ճանապարհով: Գիտության մեջ համագործակցությունը միշտ շատ ավելի արդյունավետ է լինում
Անկախ նրանից, թե դա ժառանգական է հիվանդությունԸստ Մենդելյան տեսակներից մեկի կամ պարզապես ավելի տարածված է հիվանդների հարազատների մոտ, գենետիկական ներդրումը հիվանդության մեջ բաղկացած է ընտանիքի անդամների միջև գենոտիպային տարբերություններից, որոնք կամ առաջացնում են հիվանդություն կամ փոփոխում են հիվանդության նկատմամբ զգայունությունը այս կամ այն ուղղությամբ:
Գենոմիկա, նախագծի իրականացման ընթացքում ավարտելով մարդու ԴՆԹ-ի հաջորդականության ուսումնասիրությունը, գենետիկներին տրամադրեց մարդու բոլոր գեների ամբողջական ցանկը, դրանց դիրքի և կառուցվածքի իմացությունը և տարբեր պոպուլյացիաներում հայտնաբերված մի քանի միլիոն ԴՆԹ հաջորդականության տարբերակների կատալոգ: . Այս տարբերակներից մի քանիսը շատ տարածված են, մյուսները հազվադեպ են, իսկ մյուսները հաճախականությամբ տարբերվում են տարբեր էթնիկ խմբերում:
Որոշ տարբերակներ ունեն հստակ ֆունկցիոնալ հետևանքներ, մյուսներն անկասկած չեզոք են։ Շատերի համար մարդու առողջության համար նշանակությունը անհայտ է:
Մենք նաև ուրվագծեցինք ընտրության և գենետիկ դրեյֆի դերը, որը ազդում է պոպուլյացիայի մեջ տարբեր ալելների հաճախականության վրա: Մեր MedUniver կայքի հոդվածներում մենք կքննարկենք, թե ինչպես է ժամանակի և տարածության մեջ ազդող մեյոզի գործընթացը որոշում գեների և պոլիմորֆ տեղանքների միջև կապը նրանց միջավայրի հետ:
Նախ կներկայացնենք, թե ինչ տեղեկություններ են « գենետիկական լանդշաֆտՄարդու գենոմը տրամադրվել է գենետիկական տարբերակների ժառանգության ուսումնասիրություններով, այնուհետև մենք նկարագրելու ենք հիվանդության գեների նույնականացման երկու հիմնարար մեթոդներ: Առաջին մեթոդը՝ կապի վերլուծությունը, հիմնված է ընտանիքների հարցումների վրա:
Կապի վերլուծությունունի հստակ առավելություններ ընտանեկան տոհմերի նկատմամբ՝ որոշելու հիվանդության բազմասերունդ ժառանգականությունը՝ թույլ տալով հայտնաբերել որոշակի գենոմային շրջանի հետևողական, շարունակական փոխանցումը, երբ հիվանդությունը փոխանցվում է ընտանիքում: Երկրորդ մեթոդը՝ ասոցիացիայի վերլուծությունը, հիմնված է բնակչության հարցումների վրա: Ասոցիացիայի վերլուծությունը ուղղակիորեն կախված չէ տոհմից, այլ ուսումնասիրում է որոշակի ալելի կամ ալելների հավաքածուի հաճախականության աճը կամ նվազումը պոպուլյացիայից վերցված հիվանդների ընտրանքում՝ համեմատած առողջ մարդկանց վերահսկիչ խմբի հետ:
Ասոցիացիայի վերլուծությունառավելություն ունի՝ վերլուծելով ամբողջ պոպուլյացիան՝ փնտրելու ալելներ, որոնք քիչ թե շատ հաճախ հայտնաբերվում են հիվանդների մոտ՝ համեմատած առողջ հսկիչների հետ:
Հետազոտություն կալանքԵվ ասոցիացիաներհիվանդության գեների նույնականացումը մեծ ազդեցություն է ունեցել բազմաթիվ հիվանդությունների պաթոգենեզի և պաթոֆիզիոլոգիայի ըմբռնման վրա: Այս գիտելիքը նաև հանգեցրել է կանխարգելման, բուժման և վերահսկման նոր մեթոդների մշակմանը:
Հիվանդության գեների քարտեզագրումն ունի ուղղակի կլինիկական կիրառություն՝ տրամադրելով գենի դիրքի տեղեկատվություն, որն օգտագործվում է նախածննդյան և նախասիմպտոմատիկ ախտորոշման և կրիչի հայտնաբերման համար անուղղակի կապի վերլուծության մեթոդներ մշակելու համար:
Հիվանդության գեների քարտեզագրումը չափազանց կարևոր է հիվանդության գենի նախնական որոնման համար: Քարտեզագրումը կենտրոնանում է գենոմի սահմանափակ հատվածի վրա, որպեսզի կատարի դրանում առկա բոլոր գեների քարտեզագրումը, որպեսզի մենք կարողանանք գտնել հիվանդության առաջացմանը նպաստող մուտացիաներ կամ տարբերակներ (դիրքային կլոնավորում):
Հիվանդության գենի դիրքային կլոնավորումը հնարավորություն է տալիս հիվանդությունը բնութագրելու տեղանքի տարասեռության աստիճանով, ալելային տարասեռության սպեկտրով, տարբեր պոպուլյացիաներում տարբեր պաթոգեն կամ հիվանդություն նախատրամադրող տարբերակների հաճախականությամբ, մուտացիաների ներթափանցմամբ և սպասվող հաճախականությամբ, համամասնությամբ: հիվանդության մեջ ընդհանուր գենետիկական ներդրումը, որը փոխկապակցված է տարբեր տեղաբաշխման և բնության հիվանդությունների տարբերակների հետ ռիսկի խմբի ասիմպտոմատիկ հիվանդների մոտ:
Գենների և մուտացիաների նկարագրությունըբարելավում է հիվանդությունների պաթոգենեզի ըմբռնումը և նպաստում այնպիսի ոլորտների զարգացմանը, ինչպիսիք են.
- զգայուն հատուկ ախտորոշում մուտացիայի ուղղակի հայտնաբերմամբ.
- բեռնափոխադրումների համար բնակչության սքրինինգ՝ հիվանդների կամ նրանց սերունդների մոտ հիվանդության ռիսկի խմբերը բացահայտելու համար.
- բջջային և կենդանական մոդելներ;
- դեղորայքային թերապիա հիվանդության կանխարգելման և բուժման կամ դրա առաջընթացը դանդաղեցնելու համար.
- գեների փոխարինման բուժում.

Ցանկացած Փառատոնի ժողով Սպորտային միջոցառում Շնորհանդես Հանդիպում Կոնգրես Համերգ Իրադարձություն Մրցույթներ Ցուցահանդես-կոնֆերանս...

Աշխատանք փնտրելը, երբ արդեն աշխատանք ունես, կարող է բավականին դժվար լինել, բայց դա լավագույն բաներից մեկն է, որ կարող ես...

Սթիվ Մաքքարիի աշխատանքը ներառում է բազմաթիվ պատկերային պատկերներ ֆոտոլրագրության և վավերագրական լուսանկարչության ոլորտներում: Նրա պտուղները...

Հրավիրում եմ ձեզ նայելու անսովոր ծովային նավերը, դրանք շատ բազմազան են և նման չեն իրենց մյուս գործընկերներին...
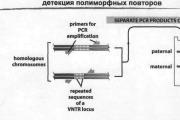
Քարտեզագրումը կոնտիգները կցելու ամենահեշտ ձևն է: Գենոմի քարտեզագրումը կարող է լինել գենետիկ և ֆիզիկական…

Որպես հիմնական մեխանիզմ, որը նախատեսված է միջին մակարդակի որակյալ մասնագետների վերապատրաստման...

Հաստատությունը գործում է համաձայն 44-FZ: 2018 թվականի գնումների պլանը կազմելիս ավելացրել ենք նոր հաստիք...

Գնումներ կատարող հաճախորդները 2011 թվականի հուլիսի 18-ի թիվ 223-FZ «Գնումների մասին» դաշնային օրենքին համապատասխան...

Իրավաբանական անձի գրանցման կարգը Մեր երկրում օրինական ձեռնարկատիրական գործունեություն իրականացնելու...

Ինձ համար սա փրկօղակ դարձավ։ Ես խրված էի խնդիրների մեջ... Ժամանակն էր փոխել կյանքս։ Նրանք ինձ օգնեցին այս հարցում...

Ինտերնետային ստարտափների բումը չի մարում. Եվ մեծ թվով մարդիկ շտապեցին զբաղվել վենչուրային ներդրումներով։ Բոլոր...

(սեղմեք բացելու համար)Աշխատեք նախատոնական օրը՝ համաձայն Արվեստի 1-ին մասի: Ռուսաստանի Դաշնության աշխատանքային օրենսգրքի 95-ը (այսուհետ՝ աշխատանքային օրենսգիրք...

Բոնուսային համակարգը յուրաքանչյուր կազմակերպությունում սահմանվում է ինքնուրույն: Այն սահմանվում է Ռուսաստանի Դաշնության աշխատանքային օրենսգրքի թիվ 114 հոդվածով,...

Կառավարիչների, մասնագետների և այլ աշխատողների պաշտոնների միասնական որակավորման գրացուցակ (ԱՄՆ), 2019թ.

Աշխատանք փնտրելը, երբ արդեն աշխատանք ունես, կարող է բավականին դժվար լինել, բայց դա լավագույն բաներից մեկն է, որ...

Սթիվ Մաքքարիի աշխատանքը ներառում է բազմաթիվ խորհրդանշական պատկերներ ֆոտոլրագրության և վավերագրական լուսանկարչության բնագավառում։